Annotated bibliography:
Acheson D, Gresack J, Risbrough V. 2012. “Hippocampal dysfunction effects on context memory: possible etiology for posttraumatic stress disorder” Neuropharmacology 62(2): 674-85
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3175276/
the hippocampus is critical for encoding memories in which a complex configuration of multiple cues is associated with the aversive event. Conversely, the hippocampus is not required for associations with discrete cues. In animal studies, if configural memory is disrupted, learning strategies using discrete cue associations predominate. These data suggest poor hippocampal function could bias the organism towards forming multiple simple cue associations during trauma, thus increasing the chances of fear responses in multiple environments (or contexts) in which these cues may be present. Here we will examine clinical and animal literature to support a theory of hippocampal dysfunction as a primary contributory factor to the etiology of PTSD
Epigenetic factors that contribute to increased stress responses in adulthood are particularly interesting as these can both be non heritable (e.g. methylation occurring due to early life stress; McGowan et al., 2009) and heritable depending upon the developmental period of epigenetic programming (e.g. in utero methylation occurring in primordial germ cells that is passed on to subsequent offspring; Dunn et al., in press)
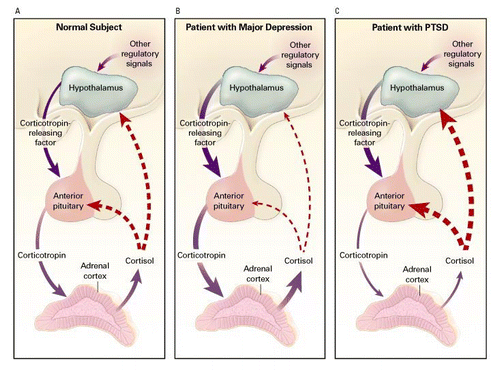
Another robust finding in neuroimaging research is reduced gray matter volume in the ACC in patients with PTSD (Yamasue et al., 2003; Corbo et al, 2005; Kitayama et al., 2006). Kasai and colleagues (2008)found that combat-exposed twins with PTSD had lower gray matter volumes in the subgenual ACC compared to their trauma-exposed non-PTSD co-twins. This pattern of finding suggests that subgenual ACC volume reductions are an acquired feature of PTSD
Pooled effect size estimates suggested significant bilateral hippocampal volume reduction in PTSD patients compared to controls. Specifically, PTSD patients had, on average, 7.2% lower right hippocampal volume and 7.0% lower left hippocampal volume. These reductions were somewhat attenuated (Right: 4.3%; Left: 4.5%) when comparing PTSD patients to trauma-exposed controls rather than control subjects without trauma exposure
In PTSD patients, studies have shown hippocampal dysfunction in the form of decreased NAA even in the absence of structural abnormalities (Freeman et al., 2006; Schuff et al., 2001). Karl and Werner (2010)conducted a recent meta-analysis of 16 total MRS studies, 12 of which provided data specific to the hippocampus. They found consistent bilateral hippocampal NAA reductions in PTSD as compared to trauma-exposed non-PTSD subjects, but not when compared to healthy controls. The authors suggested that these findings might indicate resilience, indexed by higher pre-trauma NAA levels, in the trauma-exposed non-PTSD controls relative to a mixed sample of vulnerable and non-vulnerable individuals in the healthy control groups
However when successfully encoding images that are trauma-related, PTSD subjects have been reported to exhibit reduced hippocampal BOLD responses (Hayes, LaBar, McCarthy, Selgrade, Nasser et al, in press). This study also found that reduced hippocampal activation was associated with increased arousal symptoms
Many neuropsychological studies have found deficits in verbal declarative memory in patients with PTSD (Gilbertson et al., 2006; Buckley et al., 2000; Elzinga & Bremner, 2002; Brewin, 2001; Golier & Yehuda, 1998; Bremner et al., 1993; Bermner et al., 1995; Gilbertson et al., 2001; Jenkins et al, 1998; Moradi et al., 1999; Roca & Freeman, 2001; Uddo et al., 1993; Vasterling et al., 1998; Vasterling et al., 2002; Yehuda et al., 1995; Barrett e al, 1996; Gil et al., 1990; Sachinvala et al., 2000; Golier et al, 1997), though some conflicting findings have been reported (Stein et al., 1999; Zalewski et al., 1994). Brewin and colleagues (2007) reviewed 27 studies on verbal and/or visual memory differences for emotionally neutral material between PTSD and healthy controls. The results consistently supported a small to moderate effect for poorer performance on verbal as opposed to visual memory in PTSD subjects
Recently a mutation in the estrogen response binding element of the pituitary adenylate cyclase-activating polypeptide receptor (PAC1) gene was associated with PTSD in women (Ressler et al. 2011). Interestingly this receptor is involved in hippocampal plasticity (Yang et al. 2010), but it is unknown if this mutation is linked to hippocampal function or volume in humans
expression of FKBP5, a cochaperone peptide that inhibits nuclear translocation of bound glucocorticoid receptors, is linked to risk for PTSD in subjects with early life stress (see Mehta and Binder review in this issue)
it has been hypothesized that that trauma exposure induces HPA axis dysregulation, resulting in hippocampal atrophy in vulnerable individuals (Elzinga and Bremner, 2002)
The present article proposes a mechanism through which hippocampal dysfunction may interact with traumatic experience to influence the etiology and maintenance of PTSD. Namely, an inability to adequately form conjunctive context representations may leave an individual dependent upon an elemental context representation strategy, and thus prone to respond with fear in the future presence of single elements encoded during the trauma
Adermark L, Lovinger D. 2009. “Frequency-dependent inversion of net striatal output by endocannabinoid-dependent plasticity at different synaptic inputs” J Neurosci 29(5): 1375-1380
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2744205/
Understanding how striatal neurons integrate glutamatergic and GABAergic inputs is essential for understanding the control of movement and the formation of striatal-based memories. Here we show that GABAergic synapses on striatal medium spiny neurons (MSNs) are more sensitive than glutamatergic synapses on the same cells to endocannabinoid (eCB) signaling, and that protocols that induce short-lasting cannabinoid 1 receptor (CB1R)-dependent depression at glutamatergic synapses are sufficient to induce LTD at GABAergic synapses. We also show that the frequency and duration of glutamatergic input are strong determinants of the net effect of eCB signaling, and key factors in determining if LTD has a net disinhibitory or inhibitory action in striatum. Plastic changes in net output from striatal MSNs are thus a complex function of disinhibitory and inhibitory LTD combined with other forms of synaptic plasticity such as long-term potentiation (LTP) at excitatory synapses
The striatum is the largest nucleus of the basal ganglia and has important roles in planning and execution of controlled movements (Balleine et al., 2007; Graybiel et al., 1994). The majority of synapses (~80%) are asymmetric glutamatergic synapses originating from the cortex (associative areas of the frontal and parietal lobes and motor and somatosensory cortex) and thalamus (lateral parafascicular nucleus and the central lateral nucleus) (Wilson, 2007), while the majority of neurons in the striatum (>90%) are GABAergic medium spiny neurons (MSNs) (Tepper et al., 2007).
GABAergic, presumed inhibitory, synapses are more sensitive to eCB signaling and LTD induction as compared to glutamatergic excitatory synapses in the striatum
the conditions necessary to convert CB1R-mediated short-term synaptic depression to LTD likely vary in a synapse-specific manner
Our findings indicate that with low frequency, short duration repetitive activation of glutamatergic inputs LTD of GABAergic synapses predominates; giving rise to DLL of striatal output. At higher frequencies, or longer stimulus durations, the glutamatergic synapses themselves are depressed, which would decrease output from projection neurons. The frequency of glutamatergic input is thus a strong determinant of the net effect of eCB signaling, and a key factor in determining whether LTD has a net disinhibitory or inhibitory action in striatum
Adriani W, Laviola G. 2004. “Windows of vulnerability to psychopathology and therapeutic strategy in the adolescent rodent model” Behav Pharmacol 15(5-6):341-52
http://www.ncbi.nlm.nih.gov/pubmed/15343057
Impulsive SHR animals were characterized by reduced cannabinoid CB1 receptor density in the prefrontal cortex. acute cannabinoid agonist increased levels of self-control behaviour. modulation of cannabinoid system might improve some behavioural anomalies seen in ADHD
Aharonovich E, Brooks A, Nunes E, Hasin D. 2008. “Cognitive deficits in marijuana users: effects on motivational enhancement therapy plus cognitive behavioral therapy treatment outcome” Drug Alcohol Depend 95(3): 279-283
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2429981/
Twenty marijuana-dependent outpatients were administered a neuropsychological battery at treatment entry. All patients received 12 weekly individual sessions of combined motivational enhancement therapy and cognitive behavioral therapy
Marijuana abstinence was unrelated to cognitive functioning. However, dropouts scored significantly lower than completers on measures of abstract reasoning and processing accuracy, providing initial evidence that cognitive functioning plays a role in treatment retention of adult marijuana dependent patients
participants were excluded if they 1) met DSM-IV criteria for current psychiatric disorders requiring intervention (assessed by the SCID) or substance dependence other than cannabis; 2) current use of prescription psychoactive medication 3) history of seizure disorder or head injury with loss of consciousness >1 hour, or 4) prior diagnosis of learning disability
To preclude acute intoxication at testing, patients reported their last marijuana use and submitted observed urine specimens and a breath alcohol test. Participants reporting any new illicit drug or alcohol use <7 h. before testing were rescheduled to avoid acute intoxication effects
Microcog scores among completers ranged from the population mean to about.5 SD above the mean (99.9–107.1). Microcog scores among dropouts ranged from nearly one standard deviation below the population mean to near the population mean (87.7–99.5). Dropouts had significantly lower scores on abstract reasoning, spatial processing and accuracy (), with similar trends (p<.10) for general cognitive performance and general cognitive proficiency. For example, completers scored about .5 SD above the population mean on abstract reasoning, while dropouts scored approximately .5 SD below the population mean. In contrast, WCST scores of perseverative errors and responses were near the mean and similar among completers and dropouts.
The mean proportion of negative urines did not differ between the high and low cognition groups (.18 and .17, z-statistic 0.47; p=.64)
Higher cognitive ability in the domains of mental reasoning, spatial ability and overall accuracy significantly distinguished completers from dropouts. The association between lower cognition levels at treatment entry and treatment dropout is consistent with studies in cocaine-dependent patients (Aharonovich et al., 2003; 2006) and poly-substance abusers (Fals-Stewart et al., 1994;Teichner et al., 2002)
The study was not designed to investigate differences in acute vs. residual effects of cannabis, or pre-morbid impairment vs. impairment secondary to marijuana dependence
from a treatment development viewpoint, given the high dropout early in outpatient treatment, the source of cognitive impairment may be less important than the need to tailor the initial sessions to accommodate deficits of cognitively impaired patients
Aizer A. 2013. “Juvenile incarceration, human capital and future crime: evidence from randomly assigned judges” National Bureau of Economic Research working paper no 19102
http://www.udesa.edu.ar/files/UAEconomia/Seminarios/2013/Aizer_2013.pdf
Over 130,000 juveniles are detained in the US each year with 70,000 in detention on any given day
This paper uses the incarceration tendency of randomly-assigned judges as an instrumental variable to estimate causal effects of juvenile incarceration on high school completion and adult recidivism. Estimates based on over 35,000 juvenile offenders over a ten-year period from a large urban county in the US suggest that juvenile incarceration results in large decreases in the likelihood of high school completion and large increases in the likelihood of adult incarceration
At the end of 2011, over 2.2 million people were incarcerated in the US, and an additional 4.8 million were under supervision of correctional systems (Glaze and Parks, 2012). Federal, state, and local expenditures on corrections exceed $82 billion annually, with the direct expenditures on the wider justice system totalling over $250 billion (Kennelman, 2012). Meanwhile, private expenditures that aim to prevent the externalities associated with crime are thought to be of a similar magnitude (Becker (1968). Papers and reviews include Levitt (1998, 2004); Freeman (1996); Glaeser and Sacerdote (1999); Jacob and Lefgren (2003); Di Tella and Schargrodsky , forthcoming; Lee and McCrary (2005); Lochner and Moretti (2004), among others.)
US has a juvenile corrections rate that is five times higher than the next highest country (Hazel, 2008)
In a life-cycle context, incarceration during adolescence may interrupt human and social capital accumulation at a critical moment leading to reduced future wages in the legal sector and greater criminal activity
those incarcerated as a juvenile are 39 percentage points less likely to graduate from high school and are 41 percentage points more likely to have entered adult prison by age 25 compared with other public school students from the same neighborhood. Once we include demographic controls, limit our comparison group to juveniles charged with a crime in court but not incarcerated, and instrument for incarceration, juvenile incarceration is estimated to decrease high school graduation by 13 percentage points and increase adult incarceration by 22 percentage points
the strongest results are for juveniles aged 15 and 16 – a critical period of adolescence when incarceration is most likely to end one’s high school education
In the economic model of crime originally developed by Becker (1968), criminal activity and participation in the legitimate market are substitutes
Freeman (1992) and Western and Beckett (1999). Both find that men who have been incarcerated have lower levels of employment compared with those who have not been incarcerated, controlling for an extensive set of observable characteristics
Alger B, Kim J. 2011. “Supply and demand for endocannabinoids” Trends Neurosci 34(6): 304-315
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3106144/
The endocannabinoid system in the brain primarily influences neuronal synaptic communication, and affects biological – functions including eating, anxiety, learning and memory, growth and development – via an array of actions throughout the nervous system… This review focuses on recent investigations that illuminate fundamental issues of endocannabinoid storage, release, and functional roles.
In some regions, e.g., hippocampus, the highest densities of CB1R are on axon terminals of interneurons that co-express GABA and cholecystokinin (CCK) [3][4]. In other regions, such as in the cerebellum, CB1Rs are more equally distributed on both excitatory and inhibitory terminals
Glia express CB1Rs [8] and respond to CB1R agonists by releasing glutamate and influencing synaptic transmission [9]
Endocannabinoids are directly synthesized from membrane phospholipids in response to an increase in postsynaptic intracellular calcium ([Ca2+]i) alone, or combined with activation of postsynaptic GPCRs, such as group I metabotropic glutamate receptors (mGluRs) [10][11], or M1/M3 muscarinic acetylcholine receptors (mAChRs) [12][13]
Blocking anandamide degradation reduces pain [36][37], inflammation [38], depression [39], and anxiety [40], but does not cause hypothermia, movement disorders or weight gain [40], whereas blocking 2-AG degradation induces hypothermia and hypomotility and analgesia [37]. Simultaneously blocking degradation of both mimics THC in drug discrimination tests, but blocking degradation of only one does not [34]
Construction of a 2-AG-generating system in a model cell requires heterologous expression of only mGluR5, DGLα, and the structural protein Homer 2b [64]
In the striatum, anandamide inhibits the production of 2-AG in some cells by activating TRPV1 channels, thereby decreasing glutathione levels and suppressing DGL [81]
Anandamide acting at CB1Rs seems to be responsible for chronic or constitutive endocannabinoid-mediated regulation of phenomena such as pain, anxiety and analgesia, adult neurogenesis, as well as a form of homeostatic synaptic plasticity in vitro.
Alger B. 2012. “Endocannabinoids at the synapse a decade after the dies mirabilis (29 March 2001): what we still do not know” J Physiol 590(10): 2203-2212
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3424745/
Alemany S, Arias B, Aguilera M, Villa H, Moya J, Ibanez M, Vossen H, Gasto C, Ortet G, Fananas L. 2011. “Childhood abuse, the BDNF-Val66Met polymorphism and adult psychotic-like experiences” British Journal of Psychiatry 199(1): 38-42
http://bjp.rcpsych.org/content/199/1/38
Individuals exposed to childhood abuse are more likely to report positive psychotic-like experiences. Met carriers reported more positive psychotic-like experiences when exposed to childhood abuse than did individuals carrying the Val/Val genotype. Therefore, the observed gene–environment interaction effect may be partially responsible for individual variation in response to childhood abuse.
Alisic E, Zalta A, Wesel F, Larsen S, Hafstad G, Hassanpour K, Smid G. 2014. “Rates of post-traumatic stress disorder in trauma-exposed children and adolescents: meta-analysis” BJ Psych 204:335-340
http://bjp.rcpsych.org/content/204/5/335.abstract
43 independent samples (n = 3563). Samples consisting only of participants seeking or receiving mental health treatment were excluded
The overall rate of PTSD was 15.9% (95% CI 11.5-21.5), which varied according to the type of trauma and gender. Least at risk were boys exposed to non-interpersonal trauma (8.4%, 95% CI 4.7-14.5), whereas girls exposed to interpersonal trauma showed the highest rate (32.9%, 95% CI 19.8-49.3)
Alshaarawy O, Anthony J. 2015. “Cannabis smoking and serum C-reactive protein: a quantile regressions approach based on NHANES 2005-2010” Drug Alcohol Depend 147: 203-207
http://www.drugandalcoholdependence.com/article/S0376-8716(14)01929-2/abstract
Evidence suggesting possible cannabis-attributable immunomodulation emerges at CRP levels below the median (p < 0.05)
Extending pre-clinical research on cannabis-attributable immunomodulation, this study’s CRP evidence points toward possible anti-inflammatory effects of cannabis smoking
Anda R, Felitti V, Bremner J, Walker J, Whitfield C, Perry B, Dube S, Giles W. 2006. “The enduring effects of abuse and related adverse experiences in childhood. A convergence of evidence from neurobiology and epidemiology.” Eur Arch Psychiatry Clin Neurosci 256(3): 174-86
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3232061/
For persons with ≥ 4 ACEs, the risk of panic reactions, depressed affect, anxiety, and hallucinations were increased 2.5-, 3.6-, 2.4 and 2.7-fold, respectively; smoking, alcoholism, illicit drug use, and injected drug use were increased 1.8-, 7.2-, 4.5-, and 11.1-fold; the onset of substance abuse corresponds to the time of traumatization in PTSD patients
Andel H, Jansen L, Grietens H, Knorth E, van der Gaag R. 2014. “Salivary cortisol: a possible biomarker in evaluating stress and effects of interventions in young foster children?” European Child & Adolescent Psychiatry 23(1): 3-12
http://link.springer.com/article/10.1007%2Fs00787-013-0439-1
systematic review; nine studies of salivary cortisol in foster families. All found reduced levels in relation to chronic stress related to maltreatment and neglect.
Anderson S, Bechara A, Damasio H, Tranel D, Damasio A. 1999. “Impairment of social and moral behavior related to early damage in human prefrontal cortex” Nature Neuroscience 2(11): 1032-1037
http://www.ncbi.nlm.nih.gov/pubmed/10526345
The long-term consequences of early prefrontal cortex lesions occurring before 16 months were investigated in two adults. As is the case when such damage occurs in adulthood, the two early-onset patients had severely impaired social behavior despite normal basic cognitive abilities, and showed insensitivity to future consequences of decisions, defective autonomic responses to punishment contingencies and failure to respond to behavioral interventions. Unlike adult-onset patients, however, the two patients had defective social and moral reasoning, suggesting that the acquisition of complex social conventions and moral rules had been impaired. Thus early-onset prefrontal damage resulted in a syndrome resembling psychopathy
Anderson G, Maes M. 2013. “Schizophrenia: linking prenatal infection to cytokines, the tryptophan catabolite (TRYCAT) pathway, NMDA receptor hypofunction, neurodevelopment and neuroprogression” Prog Neuropsychopharmacol Biol Psychiatry 42:5-19
http://www.ncbi.nlm.nih.gov/pubmed/22800757
In 1995, the macrophage-T lymphocyte theory of schizophrenia (Smith and Maes, 1995) considered that activated immuno-inflammatory pathways may account for the higher neurodevelopmental pathology linked with gestational infections through the detrimental effects of activated microglia, oxidative and nitrosative stress (O&NS), cytokine-induced activation of the tryptophan catabolite (TRYCAT) pathway and consequent modulation of the N-methyl d-aspartate receptor (NMDAr) and glutamate production… Accumulating data suggest a powerful role for prenatal infection, both viral and microbial, in driving an early developmental etiology to schizophrenia. Models of prenatal rodent infection show maintained activation of immuno-inflammatory pathways coupled to increased microglia activation.
Maternal infection and subsequent immuno-inflammatory responses are additionally associated with O&NS, including lowered antioxidants such as glutathione. This will contribute to alterations in neurogenesis and myelination. In such a scenario a) a genetic or epigenetic potentiation of immuno-inflammatory pathways may constitute a double hit on their own, stimulating wider immuno-inflammatory responses and thus potentiating the TRYCAT pathway and subsequent NMDAr dysfunction and neuroprogression; and b) antipsychotic-induced changes in immuno-inflammatory, TRYCAT and O&NS pathways would modulate the CNS glia-neuronal interactions that determine synaptic plasticity as well as myelin generation and maintenance
Anderson M, Rees D, Sabia J. 2013. “Medical marijuana laws and suicides by gender and age” American Journal of Public Health 104(12): 2369-2376
http://ajph.aphapublications.org/doi/abs/10.2105/AJPH.2013.301612
legalization was associated with a 10.8% (95% confidence interval [CI] = −17.1%, −3.7%) and 9.4% (95% CI = −16.1%, −2.4%) reduction in the suicide rate of men aged 20 through 29 years and 30 through 39 years, respectively
The negative relationship between legalization and suicides among young men is consistent with the hypothesis that marijuana can be used to cope with stressful life events. However, this relationship may be explained by alcohol consumption
Andersson N, Gustafsson L, Okkels N, Taha F, Cole S, Munk-Jorgensen P, Goodwin R. 2015. “Depression and the risk of autoimmune disease: a nationally representative, prospective longitudinal study” Psychological Medicine 45(16): 3559-3569
http://journals.cambridge.org/action/displayAbstract?fromPage=online&aid=10017598&fulltextType=RA&fileId=S0033291715001488
A prospective population-based study including approximately 1.1 million people was conducted using linked Danish registries. Depression and autoimmune diseases were diagnosed by physicians and documented in medical records. In total, 145 217 individuals with depression were identified between 1995 and 2012. Survival analyses were used to estimate the relative risk of autoimmune disease among those with, compared to without,
Depression was associated with a significantly increased risk of autoimmune disease [incidence rate ratio (IRR) 1.25, 95% CI 1.19–1.31], compared to those without a history of depression. Results suggest a general increased risk of autoimmune diseases following the onset of depression during first year (IRR 1.29, 95% CI 1.05–1.58), which remained elevated for the ensuing 11 years and beyond (IRR 1.53, 95% CI 1.34–1.76)
Andréasson S, Allebeck P, Engström A, Rydberg U. 1987. “Cannabis and schizophrenia. A longitudinal study of Swedish conscripts” Lancet 2(8574):1483-6
http://www.ncbi.nlm.nih.gov/pubmed/2892048
Relative risk for schizophrenia among high consumers of cannabis (use on more than fifty occasions) was 6·0
Angrilli A, Spironelli C, Elbert T, Crow T, Marano G, Stegagno L. 2009. “Schizophrenia as failure of left hemispheric dominance for the phonological component of language” PLoS One 4(2): e4507
http://www.ncbi.nlm.nih.gov/pubmed/19223971
the deficit of lateralization in the schizophrenic brain is specific for the phonological component of language
Anisman H, Merali Z, Hayley S. 2008. “Neurotransmitter, peptide and cytokine processes in relation to depressive disorder: comorbidity between depression and neurodegenerative disorders” Prog Neurobiol 85(1): 1-74
http://www.ncbi.nlm.nih.gov/pubmed/18346832/
inflammatory processes may influence stress-related illness, such as depression, and may be a common denominator for the comorbidity that exists between depression and neurological conditions, including Parkinson’s and Alzheimer’s diseases, as well as cardiovascular-related pathology
Anketell C, Dorahy M, Shannon M, Elder R, Hamilton G, Corry M, MacSherry A, Curran D, O’Rawe B. 2010. “An exploratory analysis of voice hearing in chronic PTSD: potential associated mechanisms” J Trauma Dissociation 11(1): 93-107
http://www.ncbi.nlm.nih.gov/pubmed/20063251/
Auditory hallucinations (AH) in chronic PTSD is not a rare phenomenon, (b) dissociation is significantly related to AH, and (c) dissociation may be a potential mediating mechanism for AH in PTSD.
Anselmetti S, Cavallaro R, Bechi M, Angelone S, Ermoli E, Cocchi F, Smeraldi E. 2005. “Psychopathological and neuropsychological correlates of source monitoring impairment in schizophrenia” Psychiatry Research 150: 51-59
http://www.ncbi.nlm.nih.gov/pubmed/17289157 in dropbox
Schizophrenic patients are known to show a deficit in the source monitoring function, which refers to the set of processes involved in the attribution of an origin to memories and beliefs. A failure in source monitoring was found to be associated with Schneiderian delusions in the recent literature
Recognition of self-generated items was significantly worse than control values in Schneiderian delusional patients only, while source attribution of recognized self-generated items was significantly biased towards the external sources in all delusional patients in comparison to controls. Among schizophrenic patients, source misattribution of self-generated items was significantly correlated to executive, planning performance
theory of Johnson et al. (1993) , most source monitoring decisions are made rapidly and unconsciously, using memory processes, but when percep ual and contextual information is weak , the judgment about the origins of a memory is supported by a reasoning process expect ed to involve executive functions
attribution errors of self-generated items were specifically related to the presence of active delusions. The exclusive influence of the presence of delusions and not of other positive symptoms like hallucinations or disorganization, might appear inconsistent with some previous reports (Bentall et al., 1991; Vinogradov et al., 1997; Bre´bion et al., 2000; Morrison and Haddock, 1997). However, the choice to study a sample of stabilized, an tipsychotic-responder patients may explain differences in results
Among schizophrenic patients only the executive (planning) performance (and not memory performance) was correlated to internal-external discrimination bias
might suffer from a cognitive bias consisting of a tendency to attribute “puzzling” stimuli (as self-generated items are, with weak or no perceptual and contextual information) to the previously presented more strongly contextualized source (Vinogradov, 1997)
This interpretation is consistent with data showing that patients with delusions have a “jumping to conclusion bias” (Garety, 1991)
By contrast delusional non-Schneiderian patients showed a significant performance [impairment] in comparison both to controls and non-delusional patients in the attribution of self-generated items, with a bias towards the external source.
Arain M, Khan M, Craig L, Nakanishi S. 2015. “Cannabinoid agonist rescues learning and memory after a traumatic brain injury” Ann Clin Transl Neurol 2(3): 289-294
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4369278/
Currently, there are no effective treatments for post-TBI cognitive deficits, but research has uncovered some of the pathophysiological mechanisms which include: excitotoxicity, neuroinflammation, and neurometabolic dysfunctions with an associated increase in reactive oxygen species… the cannabinoid receptor 1 (CB1R) mediates the rewarding aspects of marijuana,5 and has been shown to decrease excitotoxicity,6suppress neuroinflammation,7 and modify neurometabolism.8
administration of a CB1R agonist (ACEA) after a moderately severe experimental TBI rescued learning and memory abilities in young adult male rats
Although the CCI injury is a focal TBI with mechanical damage highly localized to the impact area, aspects of the pathological signaling cascades associated with excitotoxicity, neuroinflammation, and metabolic dysfunctions may be dispersed both ipsilateral and contralateral to the directly affected hemisphere… cannabinoid agonist treatment may have preserved learning and memory in the TBI-treated animals by protecting the intact brain tissue that was not directly damaged by the primary injury, and that the “rescued” brain areas were then able to compensate for the lesioned areas. Alternatively, and/or in parallel, the cannabinoid receptor agonist treatment could also have limited cerebral edema and neuronal cell loss,10,11 diffuse axonal damage, decreased pathological neuroinflammatory processes, or modulated metabolic processes that preserved neuronal tissues or functions.21
Ardiel E, Rankin C. 2010. “The importance of touch in development” Paediatr Child Health 15(3): 153-156
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2865952/
Developmental delay is common in children deprived of normal sensory stimulation
Hopper and Pinneau (2) found that 10 min of additional handling per day resulted in a significant reduction in regurgitation. In addition, Casler (3) reported that institutionalized infants receiving an additional 20 min of tactile stimulation per day for 10 weeks had higher scores on developmental assessments
extra mechanosensory stimulation led to superior growth and developmental performance. Although caloric consumption did not differ between the two groups, infants receiving mechanosensory stimulation averaged 47% greater weight gain per day than the unstimulated controls and were discharged an average of six days earlier. The stimulated infants also spent more time awake and active, and exhibited more mature habituation, orientation, motor and range-of-state behaviours on the Brazelton Neonatal Behavioral Assessment Scale. The positive effects appear to be persistent; when retested eight and 12 months after treatment, the stimulated infants were in a higher weight percentile group, scored better on the Bayley mental and motor assessment tests, and had a reduced incidence of neurological soft signs (minor neurological abnormalities indicating nonspecific cerebral dysfunction)
Hammett (7) reported that rats that were infrequently handled were more timid, apprehensive and high strung than rats that had been ‘petted and gentled’. They were also six times less likely to survive thyroidectomy
Gonzalez et al (8) compared the adult behaviour of maternally reared rats with those isolated in plastic cups, from postnatal days 4 to 20. Despite receiving comparable nutritional input, the pups raised in cups weighed less at weaning. Although this difference did not persist into adulthood, early deprivation did affect adult maternal and emotional behaviour. Compared with maternally reared controls, isolate-reared rats were less attentive to their own offspring, performing fewer pup retrievals and spending less time licking and crouching over pups and spending more time digging, biting the cage, hanging from the top of the cage, eating and tail chasing
full-body stroking partially rescued the behavioural deficits of isolation, with the maximally stimulated pups exhibiting maternal behaviours of durations intermediate to those of the maternally reared and minimally stimulated pups
Lovic and Fleming (9)… isolate-reared rats were hyperactive, easily distracted and less attentive to their own pups – behaviours which could be rescued by introducing licking-like stimulation with a paintbrush
the mechanism of inheritance was nongenomic: biological female offspring of low LG (licking/grooming) mothers fostered into the litter of high LG mothers displayed increased LG levels comparable with unfostered offspring of high LG mothers and vice versa… those reared by low LG mothers displayed impaired learning and memory in the Morris water maze and in object recognition (11,12), and showed substantially more fearful behaviour, as measured by a longer latency to eat food in a novel environment and decreased open-field exploration (13,10). These behavioural studies demonstrate that mechanosensory stimulation can alter the developing rat brain
Liu et al (14) showed that maternal licking altered the pup’s hypothalamic-pituitary-adrenal (HPA) stress reactivity through changes in gene expression in areas of the brain that regulate the behavioural and endocrine response to stress. Adult offspring of low LG mothers had higher HPA responses to restraint stress than those of high LG mothers
elevated exposure to stress hormones has been implicated in the development of many conditions, including visceral obesity, hypertension, diabetes, depression, anxiety, drug addiction and multiple forms of coronary artery disease
A negative feedback loop for this stress response is initiated by the binding of glucocorticoids to glucocorticoid receptors (GR) in the hippocampus. Liu et al (14) showed that as adults, the offspring of low LG mothers had decreased GR expression in the hippocampus, resulting in diminished glucocorticoid feedback sensitivity and increased CRH synthesis and release of plasma adrenocorticotropic hormone… Weaver et al (15) demonstrated that maternal care programs GR gene expression levels in the hippocampus for the lifespan of the animal via epigenetic modification of the GR gene.
administering 15 min of tactile stimulation with a small brush three times per day resulted in increased dendritic spine density in animals with mPFC lesions and increased dendritic length in animals with posterior parietal lesions. The mechanism of action appears to be the release of fibroblast growth factor-2 (FGF-2), because both FGF-2 and its receptor were upregulated in the skin and brain of stimulated rats (17)… stroking a pregnant mother rat also ameliorated the effects of future cortical lesions in her pups, as did prenatal pretreatment with FGF-2 (17)
a microscopic nematode, known as Caenorhabditis elegans, is also sensitive to touch deprivation… worms reared in isolation had a smaller body size ( and ) and a delayed onset of egg laying compared with colony worms reared in groups of 30 to 40… Rose et al (18) found that worms reared in isolation had weaker synapses than colony-reared worms, but 30 taps administered early in development were sufficient to strengthen them
Reversing the effects of early deprivation is not simple, but the importance of touch is undeniable. Finally, you need not be a worm larvae, rat pup or even human child to reap the rewards of touch. For example, employees receiving chair massages showed a significant reduction in blood pressure (21), anxiety (22) and job stress, and had increased speed and accuracy on math problems (23). Furthermore, patients with ailments ranging from burns (24) to eating disorders (25) have been shown to benefit from massage therapy, with reductions in stress hormone levels, anxiety and clinical symptoms; HIV-positive men receiving daily massages had an increased number of immune cells to combat the virus (26). To paraphrase, a kiss may just be a kiss, a sigh may just be a sigh, but a touch can change your life (or at least your nervous system)!
Arendt M, Rosenberg R, Foldager L, Perto G. 2005. “Cannabis-induced psychosis and subsequent schizophrenia-spectrum disorders: follow-up study of 535 incident cases” British Journal of Psychiatry 187: 510-515
http://bjp.rcpsych.org/content/187/6/510.full
Data on patients treated for cannabis-induced psychotic symptoms between 1994 and 1999 were extracted from the Danish Psychiatric Central Register. Those previously treated for any psychotic symptoms were excluded. The remaining 535 patients were followed for at least 3 years. In a separate analysis, the sample was compared with people referred for schizophrenia-spectrum disorders for the firsttime, but who had no history of cannabis-induced psychosis.
Schizophrenia-spectrum disorders were diagnosed in 44.5% of the sample. New psychotic episodes of any type were diagnosed in 77.2%. Male gender and young age were associated with increased risk. Development of schizophrenia-spectrum disorders was often delayed, and 47.1% of patients received a diagnosis more than a year after seeking treatment for a cannabis-induced psychosis
Cannabis-induced psychotic disorders are of great clinical and prognostic importance
535 cases (66.6%) recorded no history of psychotic symptoms and were included for further analysis.
The mean age at time of first treatment for cannabis-induced symptoms was 27.0 years (s.d.=7.7, 25th percentile=20.9, median=25.5, 75th percentile=31.2); 441 (82.4%) were male. A total of 379 patients were admitted to hospital for a median stay of 13 days (25th percentile=4, 75th percentile=29, mean=30.6) and 156 patients received out-patient treatment only.
238 people (44.5%) diagnosed with cannabis-induced psychotic symptoms later developed a schizophrenia-spectrum disorder… 77.2% experienced new psychotic episodes after index, if transient psychotic conditions and substance-induced psychoses are included. Only 15.9% remained out of psychiatric care throughout the follow-up period
The first episode of schizophrenia-spectrum disorder occurred after a substantial delay for most of the 238 patients (Fig. 1)… more than 1 year for 47.1% of the patients, and 17.2% developed such conditions more than 3 years later.
in accordance with a recent study showing that the age at onset of schizophrenia is lower among patients using cannabis (Veen et al, 2004)
The incidence of cannabis-induced psychotic disorders in Denmark was estimated to be 2.7 per 100 000 person-years. This confirms that such conditions are rare… Danish National Board of Health (2003) shows that 40.9% of all Danish citizens aged 16-24 years have used cannabis at some point in their lifetime, and that 19.7% had used the substance in the previous month
Arendt M, Rosenberg R, Fjordback L, Brandholdt J, Foldager L, Sher L, Munk-Jorgensen P. 2007. “Testing the self-medication hypothesis of depression and aggression in cannabis-dependent subjects” Psychological Medicine 37:935-945
http://www.ncbi.nlm.nih.gov/pubmed/17202003 in dropbox
119 cannabis dependent subjects, age range 16-30 (median 22) 83.2% male Subject s were excluded if there was any doubt whether cannabis dependence was the main reason for entering treatment
Nearly all subjects (92.2 %) had used cannabis more than 50 00 times and 66.4 % had used cannabis on more than 10 000 occasions. 82.4% hash most common. 48% heavily used stimulants
Depressed subjects more likely to experience sadness, depression, paranoia and anxiety and less likely to report happiness and euphoria in response to use.
Depressed subjects gave less reasons for use. The only two found more frequently in depressed than not depressed were “to give one more thoughts” and “to feel more emotions”
The majority of each group used to relax, increased pleasure and relieve depression, but depressed not moreso than non-depressed. Majority felt better while high, but a minority still felt bad. Larger minority in depression.
Conclusion is that depression does not increase the likelihood of using to relieve depression, or of experiencing relief from depression.
lowering of aggression proved to be one of the main reasons for cannabis use among subjects with difficulties controlling violent behaviour
Arendt M, Mortensen P, Rosenberg R, Pedersen C, Waltoft B. 2008. “Familial predisposition for psychiatric disorder comparison of subjects treated for cannabis-induced psychosis and schizophrenia” Arch Gen Psychiatry 65(11): 1269-1274
http://archpsyc.jamanetwork.com/article.aspx?articleid=482877
Nationwide population-based sample of all individuals born in Denmark between January 1,1955, and July 1, 1990 (N = 2 276 309)
During the 21.9 million person-years of follow-up between 1994 and 2005, 609 individuals received treatment of a cannabis-induced psychosis and 6476 received treatment of a schizophrenia spectrum disorder
rate ratios of developing cannabis-induced psychosis and schizophrenia spectrum disorder associated with predisposition to schizophrenia spectrum disorder, other psychoses, and other psychiatric disorders in first-degree relatives were of similar magnitude. However, children with a mother with schizophrenia were at a 5-fold increased risk of developing schizophrenia and a 2.5-fold increased risk of developing cannabis-induced psychosis. The risk of a schizophrenia spectrum disorder following a cannabis-induced psychosis and the timing of onset were unrelated to familial predisposition
Cannabis-induced psychosis could be an early sign of schizophrenia rather than a distinct clinical entity
almost 50% of the patients treated because of cannabis-induced psychosis in Denmark, with no history of psychosis, had a diagnosis of a schizophrenia spectrum disorder within a mean follow-up period of 5.9 years.9
the risk of developing a schizophrenia spectrum disorder was increased 3.58-fold, and the risk of developing a cannabis-induced psychosis was 4.51-fold higher in children whose father had a schizophrenia spectrum disorder compared with those whose father did not
Predisposition to psychiatric disorders other than psychosis in fathers (P = .02) was also associated with increased risk of treatment of a cannabis-induced psychosis
Despite much effort, it has been impossible to establish a symptom profile that consistently differentiates persons with cannabis-induced psychosis from those with other psychotic conditions.3,5,6,20,21 The same is true for studies comparing persons with schizophrenia who have or have not been using cannabis.4,8,10,22– 24
individuals who use cannabis or have access to the substance are at risk of having a diagnosis of cannabis-induced psychosis, although in reality they have schizophrenia
Some studies have sporadically mentioned the presence of psychopathologic findings in relatives of subjects with cannabis-induced psychosis or patients with psychoses with cannabis-positive urine screening results, but no consistent pattern has appeared.6– 8,10,26– 31 A recent study by Boydell et al4 is particularly important. These authors studied the family history of schizophrenia in 757 patients who did or did not use cannabis with onset of schizophrenia and found no difference between the groups in the percentage of patients with a positive family history of schizophrenia
However, the results clearly show that cannabis-induced psychoses do not occur randomly. Rather, the degree of hereditary predisposition in individuals who receive treatment of cannabis-induced psychosis closely mirrors that in those who develop schizophrenia with no history of cannabis-induced psychosis. The results agree with those of other studies that show that cannabis predominantly causes psychotic symptoms in those persons who are predisposed to develop psychosis or show signs of psychosis in the absence of cannabis use.7,37,39,44,45
Individuals were included in the study after having received psychiatric treatment. Consequently, they represent the more severe cases of cannabis-induced psychotic symptoms. The results may, therefore, not be generalizable to individuals who develop psychotic symptoms after cannabis use without requiring treatment or who develop psychotic symptoms that last less than 48 hours, which is required according to the ICD-10. This is important because a number of studies have shown that cannabis frequently induces short-lived psychotic symptoms both in nonpsychiatric samples and in individuals with schizophrenia.46– 50
The incidence ratio of cannabis-induced psychosis in Denmark has been estimated to be 2.7 per 100 000 person-years.9
Arnedo J, Svrakic D, Del Val C, Romero-Zaliz R, Hernandez-Cuervo H, Molecular Genetics of Schizophrenia Consortium, Fanous A, Pato M, Pato C, de Erausquin G, Cloninger C, Zwir I. 2014. “Uncovering the Hidden Risk Architecture of the Schizophrenias: Confirmation in Three Independent Genome-Wide Association Studies” Am J Psychiatry doi: 10.1176/appi.ajp.2014.14040435.
http://www.ncbi.nlm.nih.gov/pubmed/25219520
Arévalo-Martı́n Á, Vela J, Molina-Holgado E, Borrell J, Guaza C. 2003. “Therapeutic action of cannabinoids in a murine model of multiple sclerosis” Neuroscience 23(7): 2511-2516
http://www.jneurosci.org/content/23/7/2511.long
Using the Theiler’s murine encephalomyelitis virus model, we report here that treatment with the synthetic cannabinoids WIN 55,212–2, ACEA, and JWH-015 during established disease significantly improved the neurological deficits in a long-lasting way. At a histological level, cannabinoids reduced microglial activation, abrogated major histocompatibility complex class II antigen expression, and decreased the number of CD4+ infiltrating T cells in the spinal cord. Both recovery of motor function and diminution of inflammation paralleled extensive remyelination
Arévalo-Martı́n Á, Garcia-Ovejero D, Gomez O, Rubio-Araiz A, Navarro-Galve B, Guaza C, Molina-Holgado E, Molina-Holgado F. 2008. “CB2 cannabinoid receptors as an emerging target for demyelinating diseases: from neuroimmune interactions to cell replacement strategies” Br J Pharmacol 153(2): 216-225
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2219542/
Amongst the various demyelinating diseases that affect the central nervous system, those induced by an inflammatory response stand out because of their epidemiological relevance. The best known inflammatory-induced demyelinating disease is multiple sclerosis, but the immune response is a common pathogenic mechanism in many other less common pathologies
Demyelination is the loss of the myelin sheath that surrounds axons and it may affect both the central nervous system (CNS) and peripheral nervous system (PNS). Demyelinating pathologies may have a primary genetic aetiology (leukodystrophies) or may be the secondary effect of infections, vascular alterations, toxic insults or inflammatory reactions
MS is the most frequent neurological disease in young adults. In addition to myelin loss, there is also neuronal damage in MS that further contributes to the symptomatology (Ferguson et al., 1997; Mews et al., 1998; Trapp et al., 1998, 1999)
The main reason for inflammatory-mediated damage is thought to be the release of reactive oxygen and nitrogen species by immune cells, as well as that of proteases, which directly mediate cell damage (Correa et al., 2005a, 2005b). In addition, immune cells release cytotoxic/cytostatic cytokines, that not only cause damage but that may also enhance the release of more reactive species and glutamate by the cells in the surrounding tissue
Treatment of TMEV-IDD mice with a CB2 agonist reduces the infiltration of CD4+ T cells to the spinal cord (Arévalo-Martín et al., 2003)… the effect of the reduced infiltration of these cells into the CNS is beneficial as there is a decrease in the release of Th1 cytokines (interferon-γ (IFN-γ), tumour necrosis factor-α (TNF-α) or interleukin-12 (IL-12)) into the surrounding tissue, which is involved in tissue damage. It is known that cannabinoids alter the profile of cytokine expression from a Th1 to a Th2 phenotype in a CB2-dependent manner (Yuan et al., 2002)
In addition IFN-γ induces the expression of the vascular cellular adhesion molecule-1 (VCAM-1) by endothelial cells, which favours the access of activated lymphocytes to the CNS (Groveset al., 1993; Weiser et al., 2007). Therefore, reducing the release of IFN-γ by the Th1 cells could itself block the ‘calling’ signal and therefore, decrease the migration of more primed T cells to the CNS that would amplify the damage.
microglia and macrophages produce several cytokines considered to be pro-inflammatory, such as TNF-α, IL-1β or IL-12. Through CB2 receptors, cannabinoids inhibit the expression of TNF-α, IL-1β and the p40 subunit of IL-12 and IL-23 by microglia/macrophages (Klegeris et al., 2003; Correa et al., 2005a, 2005b). TNF-α causes neuronal death through an excitotoxic mechanism and recruits more lymphocytes to the CNS by upregulating the expression of VCAM-1 and intercellular adhesion molecule-1 (ICAM-1) on endothelial cells (Dobbie et al., 1999; Weiser et al., 2007)
CB2 expression is highly inducible in macrophages or microglia, and its levels in vitro depend on the local environment and the combination of inflammatory molecules (Carlisle et al., 2002; Maresz et al., 2005). In addition, in vivo CB2 is not expressed equally in all microglial populations, but rather it is predominantly present in perivascular or activated microglia (Benito et al., 2003; Nunez et al., 2004)
In [astrocytes], cannabinoids also inhibit the inflammation-induced expression of TNF-α, IL-1β and IL-6 through CB1 and CB2 receptors (Molina-Holgado et al., 1997, 1998; Ortega-Gutierrez et al., 2005)
it was recently shown that cannabinoids can prevent axonal damage in a viral model of MS, interfering with the excitotoxic component in the progression of this disease in a way that requires activation of CB2 receptors (Docagne et al., 2007)
In experimental models of demyelination there is, both neurological and histopathological, evidence of the therapeutic benefit of cannabinoids. Most data indicate that the activation of CB1 or CB2 receptors reduces deficits such as spasticity, tremor or neuropathic pain (Baker et al., 2000), whereas CB2 receptors also regulate inflammatory aspects related to disease progression (Maresz et al., 2007)
cannabinoid-induced attenuation of the inflammatory response was linked to axon remyelination. Oligodendrocyte death and the resulting destruction of myelin plays an important role in axonal degeneration, as demyelinated axons are highly vulnerable to oxidative stress and to cytokine and glutamate toxicity (Werner et al., 2001). Similarly, oligodendrocytes are sensitive to microglial-derived free-radicals and mediators of inflammation (Molina-Holgado et al., 2001; Back et al., 2002; Li et al., 2005).
once oligodendrocytes have lost their myelin membranes, they are unable to remyelinate axons even if they survive the insult (Keirstead and Blakemore, 1997)
Cannabinoids may directly enhance myelin repair by acting on oligodendrocyte progenitors, or they may act indirectly by inhibiting the immune response that might be contributing to demyelination or hampering remyelination
The synthetic cannabinoid agonists HU210 or WIN 55212-2 act on both CB1 and CB2 receptors to protect oligodendrocytes from apoptosis produced by deprivation of trophic support, a mechanism dependent on phosphoinositide 3-kinase/protein kinase B (PI3K/Akt) signalling (Molina-Holgado E et al., 2002) Moreover, cannabinoids suppress the production of inflammatory molecules by astrocytes and microglial cells including IL-1β, TNF-α and NO (Molina-Holgado et al., 1997; Molina-Holgado E et al., 2002;Puffenbarger et al., 2000; Cabral et al., 2001), as well as enhancing the release of the anti-inflammatory cytokines IL-4, IL-10, IL-6 and interleukin-1 receptor antagonist (IL-1ra) (Molina-Holgado et al., 1998,2003; Klein et al., 2000)
CB2 receptor activation could be involved in maintaining the self-renewal capacity of stem cells
Arnone D, Barrick T, Chengappa S, Mackay C, Clark C, Abou-Saleh M. 2008. “Corpus callosum damage in heavy marijuana use: Preliminary evidence from diffusion tensor tractography and tract-based spatial statistics” NeuroImage 41(3): 1067-1074
http://www.ncbi.nlm.nih.gov/pubmed/18424082 in dropbox
eleven heavy marijuana users who started using marijuana in early adolescence and eleven age matched controls
MD was significantly increased in marijuana users relative to controls in the region of the CC where white matter passes between the prefrontal lobes
Ashtari M, Avants B, Cyckowski L, Cervellione K, Roofeh D, Cook P, Gee J, Sevy S, Kumra S. 2012. “Medial temporal structures and memory functions in adolescents with heavy cannabis use.” J Psychiatr Res. 45(8): 1055-1066
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3303223/
Heavy-cannabis users showed significantly smaller volumes of the right (p< .04) and left (p< .02) hippocampus, but no significant differences in the amygdala region; smaller right correlated with a higher amount of cannabis use
Fourteen treatment-seeking/ clinically-referred male adolescents … mean age of first time cannabis use of 13.1 years (range: 9.0-15.0 years) with an average cannabis use of 5.8 joints per day from mid-to-low socio-economic, low IQ
comorbid conditions, including post-traumatic stress disorder (n = 2), attention deficit/hyperactivity disorder (n = 2), oppositional defiant/conduct disorder (n = 4), and alcohol abuse (n = 5)
Ashton J, Glass M. 2007. “The cannabinoid CB2 receptor as a target for inflammation-dependent neurodegeneration” Curr Neuropharmacol 5(2): 73-80
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2435344/
Endocannabinoids are released following brain injury and may protect against excitotoxic damage during the acute stage of injury. Brain injury also activates microglia in a secondary inflammatory phase of more widespread damage
cannabinoid CB2 receptors are up-regulated during the activation of microglia following brain injury
Taken together, studies show that CB2 is up-regulated during a process in which microglia become primed to proliferate, and then become fully reactive. In addition, CB2 activation appears to prevent or decrease microglial activation. In a rodent model of Alzheimer’s disease microglial activation was completely prevented by administration of a selective CB2 agonist. The presence of CB2 receptors in microglia in the human Alzheimer’s diseased brain suggests that CB2 may provide a novel target for a range of neuropathologies. We conclude that the administration of CB2 agonists and antagonists may differentially alter microglia-dependent neuroinflammation
It has been known since at least 1994 that the cannabinoids can be neuroprotective [4]
activation of pre-synaptic CB1 receptors is known to reduce neurotransmitter release and, hence, excitotoxicity in postsynaptic neurons [36]. Second, CB1 is involved in the regulation of vasodilation, both directly through vascular CB1 receptors and indirectly through the inhibition of the vasoconstrictor endothelin-1 [60, 71]. Third, CB1 receptors are known to regulate the release of pro-inflammatory factors such as NO and TNF-α in the acute phase of injury [18, 55]
Non-steroidal antinflammatory drugs (NSAIDS) inhibit clot formation, and increase haemorrhaging during brain injury [58]
CB2 also regulates B and T Cell differentiation, and the balance of T helper (Th1) pro-inflam-matory to Th2 anti-inflammatory cytokines [82]. In macrophages, CB2 stimulation suppresses proliferation and the release of pro-inflammatory factors such as NO, IL-12p40, and TNF-α, inhibits phagocytosis, and reduces IL-2 signalling to T-cells [9]. CB2 activation also suppresses neutrophil migration and differentiation [47], but induces natural killer cell migration [31]
CB2 regulates inflammation in a diverse range of animal models, a small sample of which includes gastro-intestinal inflammation [38], acute hindpaw inflammation [11], and pulmonary inflammation [5]. CB2-selective agonists have been particularly promising in the treatment of inflammation-induced hyperalgesia [74]
Inflammation was first suspected to be important in chronic neurodegenerative diseases such as Alzheimer’s disease (AD) early last century [14]. However, this view lost favour, largely because of the widespread belief that the blood-brain barrier provides the CNS with a privileged exemption from blood-borne leukocytes and the immune system.
The discovery that microglia are resident immune cells in the brain lead to the breakdown of this view. This was facilitated by the development of modern gene and protein expression technologies in the 1980’s and 1990’s, which lead to the discovery that many mediators of peripheral inflammation are also involved in neurodegenerative processes, including various growth factors, inflammatory chemokines and cytokines, and nitric oxide
When fully activated, microglia are amoeboid and phenotypically indistinguishable from macrophages, and carry out the various roles that macrophages perform in the periphery, including pro- and antiinflammatory functions, and antigen presentation to immune cells
2-AG induced microglial migration in a dibutyryl cAMP sensitive manner. Anandamide and palmitoylethanolamide (PEA) in combination had the same effect, though neither compound did when alone … both 2-AG and anandamide induced BV-2 microglia migration in a concentration dependent manner. Cannabinol and cannabidiol prevented the effect of 2-AG, by blocking CB2 and cannabidiol-sensitive receptors respectively
Microglia may be either neuroprotective or neurotoxic, depending upon the type and extent of exogenous or endogenous stimuli they receive and the phenotype they assume [64]. For instance, in nerve transection models of glutamate injury, microglial activity is central to the healing response [65]. By contrast, IFN-γ primed microglia then treated with LPS will adopt a phenotype adapted for defensive immunity, and hence cytotoxicity. When microglia are not reactive for defensive immune functions, however, they do not release inflammatory cytokines. Stimulation of microglia by growth factors and mitogens (e.g., M-CSF) may induce a proliferative and chemotaxic phenotype, prior to adopting either neuroprotective or neurotoxic phenotypes, depending upon other stimuli
CB2 appears to be expressed to the greatest degree when microglia are primed to proliferate. Given that 2-AG is mitogenic for this microglial phenotype, and that cannabinoids induce microglial migration and could be critical to chemotaxis, it is possible that CB2 regulates not only the cytotoxic properties of activated microglia, but also the neuroprotective properties of microglia. To our knowledge, this has yet to be determined
A further possibility is that release of the immune suppressive cytokine TGF-β is regulated in microglia by CB2. TGF-β is known to play a critical role in the regulation of TNF-α and IL-1 release from microglia [67] as well as in the induction of neuronal proliferation. In addition, Δ9-THC stimulates TGF-β production in peripheral blood lymphocytes via CB2, and TGF-β regulates lymphocyte CB2 receptor expression [22]
It has been known for several years that cannabinoids stimulate neurogenesis in the adult brain [30]. Recently, Palazuelos et al. [54] reported that CB2 receptors are highly expressed in neural progenitors and immature neurons in vitro and in vivo. Stimulation of CB2 in vitro induced neural progenitor cell proliferation and the formation of neurospheres
cannabinoids acting through the CB2 receptor inhibit the release of pro-inflammatory and cytotoxic factors such as IL-1, NO and TNF-α in microglia previously activated for defensive immunity
CB2 receptor activation appears to have benign effects on microglia potentially primed for adaptive immunity and neuroprotection, to block differentiation of microglia into a neurotoxic phenotype, and to inhibit the release of neurotoxic factors when microglia are activated. In addition, CB2 stimulation is pro-neurogenic in areas of adult neurogenesis
In bone tissue, CB2 receptors stimulate osteoblast function and inhibit osteoclasts, leading to increased bone thickness [49]. Potentially useful for osteoporosis, this also has the promise to help control key mechanisms involved in the generation of pain in bone cancer [27]
Auer R, Vittinghoff E, Yaffe K, Kunzi A, Kertesz S, Levine D, Albanese E, Whitmer R, Jacobs D, Sidney S, Glymour M, Pletcher M. 2016. “Association between lifetime marijuana use and cognitive function in middle age The Coronary Artery Risk Development in Young Adults (CARDIA) Study” JAMA Intern Med doi:10.1001/jamainternmed.2015.7841
http://www.ncbi.nlm.nih.gov/pubmed/26831916 add to dropbox
a cohort of 5115 black and white men and women aged 18 to 30 years at baseline from March 25, 1985, to June 7, 1986 (year 0), and followed up over 25 years from June 7, 1986, to August 31, 2011… Linear regression was used to adjust for demographic factors, cardiovascular risk factors, tobacco smoking, use of alcohol and illicit drugs, physical activity, depression, and results of the mirror star tracing test (a measure of cognitivefunction) at year 2
Among 3385 participants with cognitive function measurements at the year 25 visit, 2852 (84.3%) reported past marijuana use, but only 392 (11.6%) continued to use marijuana into middle age. Current use of marijuana was associated with worse verbal memory and processing speed; cumulative lifetime exposure was associated with worse performance in all 3 domains of cognitive function. After excluding current users and adjusting for potential confounders, cumulative lifetime exposure to marijuana remained significantly associated with worse verbal memory. For each 5 years of past exposure, verbal memory was 0.13 standardized units lower (95% CI, −0.24 to −0.02; P = .02), corresponding to a mean of 1 of 2 participants remembering 1 word fewer from a list of 15 words for every 5 years of use. After adjustment, we found no associations with lower executive function (–0.03 [95% CI, −0.12 to 0.07]; P = .56) or processing speed (–0.04 [95% CI, −0.16 to 0.08]; P = .51)
he attenuation of the association be-tween marijuanaexposureandall3measuresof cognitive function was seen mostly after adjustment for race and sex strata and
educationallevel.Sensitivityanalysesdemonstratednoevidence of significant interactions by race or sex
In the context of cognitive decline after stroke, Levine et al 34 used a 0.5-SD cutoff for defining a clinically meaning-ful decline in global cognition. The point estimate for verbal memory in our study for those with 5 marijuana-years of ex-posure (0.13 standardized units; 95% CI, −0.24 to −0.02) is of lesser magnitude than the decline found in the study by Levine et al and the confidence interval excludes the 0.5-SD cutoff
we cannot rule out reverse causation as an explanation for our results… factors significantly associated with marijuana use could confound the association
Aupperle R, Melrose A, Stein M, Paulus M. 2012. “Executive function and PTSD: disengaging from trauma” Neuropharmacology 62: 686-694
http://www.ncbi.nlm.nih.gov/pubmed/21349277 in dropbox
estimated 50-60% of people will experience a serious trauma as a result of combat, sexual assault, major accidents, or other real-life horrors at some point in their lives (Kessler et al., 1995). However, only 5-10% of people are estimated to develop symptoms qualifying them for diagnosis of posttraumatic stress disorder (PTSD)
William James, in The Principles of Psychology, defined attention as “the taking possession by the mind, in clear and vivid form, of one out of what seem several simultaneously possible objects or trains of thought” (James, 1890 ). He went on to say that “It implies withdrawal from some things in order to deal effectively with others. ”
1) attention, or the voluntary allocation of processing resources or focusing of one ’s mind on a particular stimulus within the environment, 2) working memory, or the active maintenance and manipulation of information in one’s mind over a short period of time, 3) sustained attention , or the maintenance of attention on one set of stimuli or a task for a prolonged period, 4) inhibitory function, involving the inhibition of automatic responses to maintain goal-directed behavior, 5) flexibility /switching, or the ability to switch between two different task s or strategies, and 6) planning ,or the ability to develop and implement strategic behaviors to obtain a future goal
attentional modification programs may be beneficial in the treatment of anxiety disorders (Amir et al., 2009a; Li et al., 2008; Schmidt et al., 2009; Amir et al., 2009b; Najmi and Amir, 2010; Amir et al., 2008). This suggests that research related to attention and working memory function may not only increase our understanding of PTSD, but may also lead to more effective treatments for these patients
usually impossible to determine whether any observed cognitive dysfunctions represent pre-trauma risk and resiliency factors or if they represent responses to the experience of trauma or PTSD
pre-trauma performances on immediate and delayed verbal recall (California Verbal Learning Test [CVLT]) (Delis et al., 1988), working memory (digit span backward), visuomotor speed (Symbol Digit Modalities Test [SDMT]) ( Smith, 1982), and verbal intelligence (National Adult Reading Test [NART]) ( Nelson, 1982; Nelson and Willison, 1991) was negatively related to post-trauma PTSD re-experiencing and arousal symptoms
Measures of overall IQ, verbal memory (immediate and delayed recall), attention (digit span) (Wechsler, 1987), and executive function (Wisconsin Card-Sorting Test [WCST]) ( Heaton, 1981) performance was decreased not only for the PTSD group, but also their twins, compared to the non-PTSD group and their twins. These results provide further support that lower pre-trauma cognitive functioning particularly in domains of attention, executive function, and memory may serve as a risk factor for the development of PTSD
cognitive function (e.g., learning and memory) found to correlate with PTSD severity above and beyond that accounted for by premorbid IQ (Vasterling et al., 2002; Gilbertson et al., 2001)
A decrease in specific cognitive functions pre-trauma may not only influence the development of PTSD, but may itself be exacerbated by the experience of trauma
Decreased performance on measures of auditory attention and working memory have been found in combat- and sexual assault- related PTSD when compared to victims without PTSD and non-trauma controls (Samuelson et al., 2006; Brandes et al., 2002; Gilbertson et al., 2001; Vasterling et al., 1998, 2002; Marmar et al., 2006; Lagarde et al., 2010; Gilbertson et al., 20 01; Jenkins et al., 2000), and these deficits have been reported to correlate with PTSD symptom severity (Burriss et al., 20 08)
it is unclear whether PTSD is associated with primary problems in attention and working memory, or whether the inconsistent findings are due to difficulties coping with and inhibiting unintentional “distracters”, such as internal (e.g., emotions, cognitions) or external stimuli (e.g., environmental sounds and sights; stimuli presented in previous tasks)
Studies have repeatedly found PTSD patients to exhibit impaired performance (e.g., increased omissions, commissions, or reaction time) in auditory and visual sustained attention (Vasterling et al., 1998; McFarlane et al., 1993; Wu et al., 2010; Shucard et al., 20 08; Jenkins et al., 20 0 0; Vasterling et al., 20 02 ); except see (Golier et al., 1997 ), and the number of correct hits has
been reported to negatively correlate with PTSD symptom severity (Vasterling et al., 20 02 ).
Impaired performance on the color-word Stroop has been reported for various PTSD populations, though it is unclear whether such a deficit is specific to PTSD or a more general impairment across psychiatric disorders (Lagarde et al., 2010; Litz et al., 1996). Interestingly, several studies have also reported PTSD to be associated with increased intrusions during memory recall (Vasterling et al., 1998; Lindauer et al., 2006)
Heightened arousal and re-experiencing symptoms could create more distracters when an individual is attempting to concentrate on the task at hand, thereby interrupting working memory, sustained attention, and inhibitory functions. However, it is also possible that primary inhibitory dysfunction could result not only in decreased performance on cognitive task s, but also impaired ability to inhibit emotional memories and physiological arousal in response to triggers
Neuropsychological research therefore seems to provide inconsistent support for impairment in speed-reliant, attentional switching, but indicates that planning, rule-learning, and untimed strategy switching, may be mostly spared in PTSD
recall of emotional autobiographical events does not influence working memory function any more for PTSD patients than controls
Two studies have been conducted thus far to examine decision making in PTSD patients, both of which found PTSD to be associated with an increase in the number of trials needed to learn optimal patterns of responding (Sailer et al., 20 08; Koenen et al., 20 01)
hyperactivation within limbic regions (particularly amygdala and insula) and hypoactivation of prefrontal regions, including anterior cingulate (ACC; including both rostral and dorsal) and ventromedial prefrontal cortex (vMPFC)
The lateral PFC specifically has been implicated in response inhibition whether it be emotional or non-emotional contexts (Compton et al., 2003; Bledowski et al., 2010). However, the ACC may have some specialization in this regard, as more ventral regions are thought to be primarily involved in inhibition of responses to emotional stimuli, while more dorsal regions are thought to be involved in the inhibition of neutral information (Whalen et al., 1998; Bush et al., 1998; Mohanty et al., 2007; Yamasaki et al., 2002; Fichtenholtz et al., 2004)
PTSD has been associated with increased activation in dorsal ACC and other PFC regions during an auditory oddball task ( Bryant et al., 2005). However, during the go-nogo task , PTSD patients exhibited reduced activation in the inferior frontal and ventral and dorsal lateral PFC, as well the medial OFC ( Falconer et al., 2008 )
PTSD may be associated with hyperactivation of prefrontal areas in response to simple sustained attention tasks, but relative hypoactivation during tasks involving inhibition or “updating” . The former could re flect the hypervigilance and enhanced attention towards“triggers ” associated with PTSD, while the latter could relate to decreased ability to control or inhibit these attentional resources
Attention modification has been effective in reducing symptoms in social anxiety (Li et al., 2008; Schmidt et al., 2009; Amir et al., 2008), generalized anxiety (Amir et al., 2009b ), and sub-clinical obsessive-compulsive disorder (Najmi and Amir, 2010)
if you can ’t inhibit it avoid it
Avitsur R, Stark J, Sheridan J. 2001. “Social stress induces glucocorticoid resistance in subordinate animals” Horm Behav 39(4): 247-57
http://www.ncbi.nlm.nih.gov/pubmed/11374910/
elevated levels of serum corticosterone and nerve growth factor (NGF). Repeated exposure to an intruder induced a state of glucocorticoid resistance in peripheral immune cells
it may be that the development of glucocorticoid resistance is an adaptive mechanism that allows the inflammatory component of wound healing to occur in the presence of high levels of corticosterone
Azad S, Monory K, Marsicano G, Cravatt B, Lutz B, Zieglgansberg W, Rammes G. 2004. “Circuitry for associative plasticity in the amygdala involves endocannabinoid signaling” J Neurosci 24(44):9953-61
http://www.ncbi.nlm.nih.gov/pubmed/15525780
Endocannabinoids are crucial for the extinction of aversive memories. 100 pulses at 1 Hz to afferents in lateral amygdala releases anandamide postsynaptically from neurons in basolateral amygdala of mice in vitro, inducing long-term depression (LTDi) of GABA, which increases amplitude of excitatory currents in principal neurons of central nucleus. LTDi abolished by CB1 antagonist and in receptor-deficient animals. Enhanced in animals lacking FAAH.
could provide a prerequisite for extinction by formation of new memory
Bäckhed F, Roswall J, Peng Y, Feng Q, Jia H, Kovatcheva-Datchary P, Li Y, Xia Y, Xie H, Zhong H, Khan M, Zhang J, Li J, Xiao L, Al-Aama J, Zhang D, Lee Y, Kotowska D, Colding C, Tremaroli V, Yin Y, Bergman S, Xu X, Madsen L, Kristiansen K, Dahlgren J, Wang J. 2015. “Dynamics and stabilization of the human gut microbiome during the first year of life” Cell Host & Microbe 17: 690-703
http://www.ncbi.nlm.nih.gov/pubmed/25974306 add to dropbox
In contrast to vaginally delivered infants, the gut microbiota of infant s delivered by C-section showed significantly less resemblance to their mothers. Nutrition had a major impact on early microbiota composition and function, with cessation of breast-feeding, rather than introduction of solid food, being required for maturation into an adult-like microbiota
Compared with vaginally born infants, the C-section fecal microbiome was enriched in MetaOTUs such as Enterobacter hormaechei/E. cancerogenus, Haemophilus parainfluenzae/H.aegyptius/H. influenzae/H. haemolyticus, Staphylococcus saprophyticus/S. lugdunensis/S. auereus, Streptococcus australis and Veillonella dispar/V. parvula, indicating that skin and oral microbes, but also baceria from the surrounding environment during delivery, were the first colonizersin these infants. In contrast, the gut microbiota of vaginally delivered newborns were enriched in microbes from the genera Bacteroides, Bifidobacterium, Parabacteroides, Escherichia/Shigella, which were also the most abundant members of the newborns’ gut microbiota. … The difference between delivery modes gradually decreased at 4 months and then 12 months of age, but the C-section infants remained more heterogeneous compared to the vaginally born infants.
most of the early colonizers of the newborn gut originate from the mother and the mode of birth is an important factor shaping the gut microbiota of term infants in early life
The gut microbiome is an important producer of vitamins. All newborns in Sweden receive prophylactic vitamin K injections to avoid classic hemorrhagic disease. We observed enriched levels of genes for vitamin K2 (menaquinone) synthesis in newborns, which correlated with the high abundance of Bacteroides and Escherichia/Shigella, known vitamin K2 producers. Vitamin K2 is important for bone and heart health, and the microbime was recently described to modulate bone homeostasis. Metabolism of retinol was also most enriched in the newborns, with implications in several essential developmental processes such as vision, bone and teeth. Vitamins from the so-called B complex are needed for the body to convert nutrients into glucose and produce energy. Folate (vitamin B9) is one of the essential B vitamins involved in DNA snthesis and repair. Folate biosynthetic genes were significantly enriched in newborns. Genes for pyridoxal (vintamin B6) and biotin (vitamin B7) synthesis were also significantly enriched in newborns. In contrast, thiamine, pantothenate and cobalamin (vitamins B1, B5, and B12, respectively) biosynthetic genes increased with age, consistent with a previous study. However, modules for vitamin B12 transport system were strongly increased in the newborn metagenome, but decreased with age. Similarly, transporters for iron, hemin and heme, which are linked to vitamin B12 synthesis and important for iron metabolism, were also increased in the microbiome of newborns.
Exclusively breast fed infants (at 4 months) had increased levels of taxa that are used as probiotics such as L. johnsonii/L.gasseri, L. paracasei/L. casei, and B. longum. Four-month-old formulat-fed infants had elevated levels of Clostridium difficile, Granulicatella adiacens, Citrobacer spp., Enterobacter cloacae, Bilophila wadsworthia, in agreement with previous studies… formula fed infants were enriched in functions found in the adult microbiome, such as bile acid biosynthesis and methanogenesis. According to the CAZy database, formula-fed infants exhibited an overrepresentation of GH86, GH116, PL1, and PL2, which are beta-agarase or beta-porphyranase and pectate lyase. In contrast, the microbiome of infants that were exclusively breast-fed had higher levels of KO modules involved in oxidative phosphorylation and synthesis of B vitamins such as riboflavin, tetrahydrofolate, and biotin and GH119.
… Yatsunenko 2012 authors concluded that the infant microbiome gradually matures into an adult-like structure until the age of 3 years. However, this might be more based on the gut microbiota in the Malawian and Amerindian children, as the 1-year old infants from the UNited States were already as close to the adults as teenagers. In our Swedish cohort, infants at 12 months were more similar to their mothers than were newborn infants or newborns at 4 months. However, differences remained in the gut microbiome between the 1-year-old children and the mothers both compositionally and functionally, awaiting further maturation.
Our results underscore the role of the gut microbiota for the production of essential amino acids and vitamins for the growing infant. While the infant gut microbiota acquired significant capacity to produce amino acids and vitamins after 4 months of life, the increase in transporters capacity indicates that the newborn’s microbiome is poised to the upcoming change in the intestinal environment and progression to a mature profile. Intriguingly, considering evidence that the gut microbiota may affect behavior, many functions of the developing gut microbiome linked to the metabolism of vitamins, iron, and amino acids are also required for normal brain development, thus adding to the possibility that the gut microbiota might affect behavior.
Baker S, Yücel M, Fornito A, Allen N, Lubman D. 2013. “A systematic review of diffusion weighted MRI studies of white matter microstructure in adolescent substance users” Neuroscience and Biobehavioral Reviews 37: 1713-1723
http://www.ncbi.nlm.nih.gov/pubmed/23845745 in dropbox
White matter alterations may reflect risk for, rather than result from, substance use.
criticism of DW-MRI a finding of lower FA within a voxel may reflect a more complex fibre tract arrangement and is not necessarily indicative of poorer white matter microstructure (Jeurissen et al., 2012) (Jones et al., 2012)
Bachhuber M, Saloner B, Cunningham C, Barry C. 2014. “Medical cannabis laws and opioid analgesic overdose mortality in the United States, 1999-2010” JAMA Intern Med 174(10): 1668-1673
http://archinte.jamanetwork.com/article.aspx?articleid=1898878
States with medical cannabis laws had a 24.8% lower mean annual opioid overdose mortality rate which strengthened over time
year 1 (−19.9%), year 2 (−25.2%), year 3 (−23.6%), year 4 (−20.2%), year 5 (−33.7%), and year 6 (−33.3%)
Bakker A, Krauss G, Albert M, Speck C, Jones L, Stark C, Yassa M, Bassett A, Gallagher M. 2012. “Reduction of hippocampal hyperactivity improves cognition in amnestic mild cognitive impairment” Neuron 74(3): 467-474
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3351697/
Ballard M, Bedi G, de Wit H. 2012. “Effects of delta-9-tetrahydrocannabinol on evaluation of emotional images” J Psychopharmacol 26(10): 1289-1298
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3664416/
THC significantly impaired recognition of facial fear and anger, but it only marginally impaired recognition of sadness and happiness. The drug did not consistently affect ratings of emotional scenes. THC’ effects on emotional evaluation were not clearly related to its mood-altering effects. These results support our previous work, and show that THC reduces perception of facial threat. Nevertheless, THC does not appear to positively bias evaluation of emotional stimuli in general
Bambico F, Duranti A, Tontini A, Tarzia G, Gobbi G. 2009. “Endocannabinoids in the treatment of mood disorders: evidence from animal models” Curr Pharm Des 15(14): 1623-46
http://www.ncbi.nlm.nih.gov/pubmed/19442178
Among all mental disorders, major depression has the highest rate of prevalence and incidence of morbidity. Currently available antidepressant therapies have limited efficacies; consequently, research on new drugs for the treatment of mood disorders has become increasingly critical. Recent preclinical evidences that cannabinoid agonists and endocannabinoid enhancers, such as the fatty acid amide hydrolase (FAAH) inhibitors, can impact mood regulation have opened a new line of research in antidepressant drug discovery
cannabinoid agonists and endocannabinoid enhancers increase serotonin and noradrenergic neuronal firing activity, increase serotonin release in the hippocampus, as well as promote neurogenesis. Since cannabinoid-derived drugs potentiate monoaminergic neurotransmission and hippocampal neurogenesis through distinct pathways compared to classical antidepressants, they may represent an alternative drug class in the pharmacotherapy of mood and other neuropsychiatric disorders
Barnes G, Barnes M, Patton D. 2005. “Prevalence and predictors of “heavy” marijuana use in a Canadian youth sample” Subst Use Misuse 40(12): 1849-63
http://www.ncbi.nlm.nih.gov/pubmed/16419560
high scores on father’s alcoholism and family life stress, and low scores on family cohesion were associated with once a week (12.6% of the sample) versus less cannabis use in sample of youth aged 14–25.
Barkus E, Lewis S. 2008. “Schizotypy and psychosis-like experiences from recreational cannabis in a non-clinical sample” Psychol Med 38(9): 1267-76
http://www.ncbi.nlm.nih.gov/pubmed/18205966
schizotypy was associated with increased psychosis-like experiences and after-effects, but also found that high-scoring schizotypes reported more pleasurable experiences when smoking cannabis. Using new subscales derived from principal components analysis (PCA), we found that the psychosis-like items were most related to varying rates of schizotypy both during the immediate use of cannabis and in the after-effects of cannabis use. High-scoring schizotypes who used cannabis experienced more psychosis-like symptoms during and after use
cannabis use may reveal an underlying vulnerability to psychosis in those with high schizotypal traits
Barkus E, Morrison P, Vuletic D, Dickson J, Ell P, Pilowsky L, Brenneisen R, Holt D, Powell J, Kapur S, Murray R. 2011. “Does intravenous Δ9-tetrahydrocannabinol increase dopamine release? A SPET study” J Psychopharmacol 25(11): 1462-8
http://www.ncbi.nlm.nih.gov/pubmed/20851843
there were no significant differences in binding index in the caudate or putamen under THC compared to placebo conditions. Positive psychotic symptoms and DA release were unrelated. THC did not lead to a significant increase in DA release even though the dose was sufficient for participants to have psychotic symptoms
Batalla A, Bhattacharyya S, Yücel M, Fusar-Poli P, Crippa J, Nogue S, Torrens M, Pujol J, Farre M, Martin-Santos R. 2013. “Structural and Functional Imaging Studies in Chronic Cannabis Users: A Systematic Review of Adolescent and Adult Findings.” PLOS ONE doi: 10.1371/journal.pone.0055821
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0055821
Of the seven studies comparing global brain volume measures between chronic cannabis users and healthy controls, there was only one study reporting significant differences
Block et al. (2000) [157] described reduced bilateral rCBF in the posterior cerebellum and ventral prefrontal cortex but increased rCBF in the anterior cingulate cortex in users. Lundqvist et al. (2001) [158] found a trend of lower global CBF in cannabis users, reduced rCBF in the right prefrontal and superior frontal cortex. Sevy et al. (2008)[159] reported lower glucose metabolism in the right orbitofrontal cortex, putamen bilaterally and precuneus in chronic cannabis users.
Chang et al. (2006) [169] used fMRI to compare a visual-attention task in current and abstinent cannabis users with healthy controls. normal task performance, both active and abstinent chronic cannabis users … decreased activation in the right prefrontal, medial and dorsal parietal cortices and medial cerebellar regions but greater activation in regions throughout the frontal, posterior parietal, occipital and cerebellum. … early age of onset and estimated cumulative cannabis lifetime exposure were both associated with reduced activation in the right prefrontal cortex and medial cerebellum.
Harding et al. (2012) [171] demonstrated long-term heavy cannabis associated with increased functional connectivity between several frontal cortex regions and the occipitoparietal cortex using the Multi-Source Interference Task (MSIT). No differences in behavioural performance were evident between groups.
Gruber et al. (2005) [151] lower anterior cingulate activity and higher mid-cingulate and bilateral dorsolateral prefrontal cortex activity in current cannabis users relative to healthy controls, focal increased activity in right dorsolateral prefrontal cortex. … Eldreth et al. (2004) [166] found in abstinent cannabis users a reduced anterior cingulate activation using H215O-PET during modified Stroop test; reduced dorsolateral prefrontal cortex activation, greater activation in the hippocampus bilaterally
cannabis users showed enhanced reward anticipation activity in the middle temporal gyrus bilaterally, right cuneus and right parahippocampal gyrus. When compared to tobacco-smokers, cannabis users showed reduced anticipation activity in the same areas, with the exception of the nucleus accumbens bilaterally, the right medial frontal gyrus and the left cingulated gyrus, indicating that anticipation activity in these regions may be attenuated by both cannabis and nicotine [176]
cannabis users demonstrated relatively lower anterior cingulate and amygdalar activity during the presentation of masked angry stimuli sets relative to the control group, who showed relatively higher activation within these regions. In contrast, cannabis users demonstrated a larger pattern of activation during the presentation of masked happy faces within the cingulate as compared to controls, with no increase in amygdalar activation [170]. Furthermore, the total number of smoking episodes per week was positively associated with cingulate activity during the viewing of masked angry faces and positively associated with amygdalar activity during the viewing of masked happy faces [170]. Finally, overall cannabinoid level was positively related to cingulate activity during the viewing of masked happy faces [170]. … suggest a different way of processing emotional information between groups [170].
In a group of abstinent adolescent cannabis users, Padula et al. (2007) [179] described increased activity in the left temporal gyrus and anterior cingulate cortex but lower activity in right temporal gyrus, thalamus, pulvinar and left parahippocampal gyrus related to higher scores on the task, while the reverse pattern was found in the controls. This may suggest that cannabis users employed more of a verbal strategy to achieve the same level of task performance as the controls [179]. Additionally, cannabis users demonstrated greater performance-related activation in the right basal ganglia, precuneus, postcentral gyrus and bilateral superior parietal lobe [179], again suggesting a compensatory neural effort
groups performed at a similar level; recent users showed greater activity in the medial and left superior prefrontal cortices and bilateral insula while abstinent users demonstrated an increased response in the right precentral gyrus [181]
…in the only study where the chronic cannabis users were not abstinent…
studies that merely compare those subjects exposed to an environmental factor from those that are not, are likely to promote interpretation biases whereby study findings, irrespective of their direction, tend to be interpreted as detrimental
Battisti R, Roodenrys S, Johnstone S, Respondek C, Hermens D, Solowij N. 2010. “Chronic use of cannabis and poor neural efficiency in verbal memory ability” Psychopharmacology 209: 319-330
http://www.ncbi.nlm.nih.gov/pubmed/20217055 — add to dropbox
The SME (subsequent memory effect) is predictive of recall outcome and originates in structures that are dense with cannabinoid receptors (hippocampus and parahippocampus). The SME and performance on a verbal memory task were compared between 24 cannabis users (mean 17 years of near daily use) in the unintoxicated state and 24 non-using controls
Cannabis users showed poorer recall and altered patterns of SME activation: specifically, attenuation of the negative N4 and an increase in the late positive component. Duration of use and age of initial use correlated significantly with SME amplitudes. A longer history of use also correlated with greater recall that was related to N4 expression
In studies of acute administration, Roser et al (2008) have recently shown that THC modulates the amplitude of teh mismatch negativity component, an early index of sensory memory; Leweke et al (1998) found enhanced positivity from 250 ms onwards following the presentation of previously learned words in an emotional word recognition task; while Ilan et al (2004, 2005) found attenuated ERP amplitudes, including a reduced N400, in spatial working memory and word recognition tasks. In studies of chronic users during the unintoxicated state, Patrick et al (1999) showed a reduced P50 auditory gating response, and Rentzsch et al (2007) determined that this P50 deficit was also evident in 28 day abstinent chronic cannabis users and correlated with the duration of cannabis use. Kempel et al (2003) found evidence of altered selective attentional proceses indexed by the N2 and P3 components, which were associated with early onset cannabis use. Skosnik et al found larger P300 amplitudes to unpleasant trait words in an affect processing task in one study (Skosnik et al 2008) and altered steady-state visual-evoked potentials in another (Skosnik et al 2006). Wolfling et al (2008) found a larger late positivity in visual ERPs to drug=related cues. Solowij et al (1991, 1995b) and Solowij (1998) observed differential effects of duration versus frequency of cannabis use in a selective attention task. Frontal processing negativity to irrelevant stimuli increased with the duration of exposure to cannabis in current users and in ex-cannabis users (mean abstinence, 2 years), suggesting persistent chagnes in the brain. P300 latency increased with increasing frequency of use in current users only, suggesting a short-term effect potentially associated with a buildup of cannabinoid residues in the brain.
Inclusion required being between the ages of 18-55 and no history of diagnosis with any psychotic disorder or neurological injury. Required to abstain from use for at least 12 h prior to the test session
Information collected included age, handedness, educational, medical, and mental health (both personal and family) histories, as well as detailed questioning regarding any type of susbstance use (including alcohol and tobacco)
6.5 drinks per week in cannabis group, 2.3 in control. 101.3 WAIS-III-R FSIQ, 106.2 for control. 75% of cannabis users were using 25 days or more per month. Median 30. 435 “cones” per month (one third of a joint)
There were more tobacco smokers within the cannabis-using group, and all but four users mixed their cannabis with tobacco.
Cannabis users recealled significantly fewer words than controls across all word lists presented vs controls. As a proportion, this represents an average reduction of 13.5% in correct recall for users in comparison to controls. The mean words recalled per list by users was 6.87, and for controls, 7.94. Percentage of CR words also correlated marginally significantly with duration of regular cannabis use. A longer history of regular use was associated with a greater number of CR words
overall shift to greater positivity (ie, reduced N4 and increased LPC) among users… users showing a shorter N4 latency than controls (mean difference = 13.88 ms)
Groups did not differe in the nature of the primary encoding strategy used, although controls appeared more likely to use visual imagery and less likely to use rote learning stategies than users, and both groups relied heavily upon categorization… user N4, relative to controls, is attenuated particularly on CR trials frontally and on NR trials parietally… User N4 was greater within the left compared to the right sites, whereas the difference was minimal for controls. This pattern varied significantly with condition. Users had greater overall variability between sites with this particularly evident on NR trials. This pattern was not evident for controls.
Controls showed increased N4 at Fz relative to the lateral sites, with this enhancement of N4 at Fz absent in users. A pattern of reduced N4 for the parietal right (P4) relative to the left hemisphere (P3) was present for users but not controls
Controls had a single correlation only with age and N4 amplitude on correct trials, whereas users had significant correlations for age, IQ, and percentage of CR words with N4 amplitude on both trial types. Higher scores on these variables were related to attenuated N4 (ie, more positive as this is a negative-going component). Positive correlations were observed between N4 amplitudes and age of first cannabis use and age of regular use, and with duration of use, although there wer no observed relationships with quantity of exposure per month, or with levels of cannabinoid metabolites or measures of recency of use or withdrawal scores. Controlling for age resulted in a loss of significance for cannabis usage history variables with N4. However, controlling for duration since first use and duration of regular use on CR and NR trials, respectively, resulted in a loss of significance for the N4 correlation with age, thus indicating relationships to be jointly explainable by age and usage variables. There were no significant correlations with N4 latency or the LPC.
An overall shift to positivity and changes to variability within regions was observed for users on both SME components of interests , with N4 earlier and attenuated and LPC increased. Altered activation of the N4 occurred on CR versus NR trials between the two groups, with a longer history of cannabis use related to attenuated N4. … users showed significantly poorer performance in verbal memory, although poorer word recall performance appeared to correlate with a shorter history of regular use. … Notably, the first 350 ms of the ERPs were similar; however, the critical N4-LPC window showed marked differences across groups and conditions. In particular, the overall shift to greater positivity may be due to N4 peaking earlier for users and interpreted as a general failure to efficiently activate memory-related brain structures
Bava S, Frank L, McQueeny T, Schweinsburg B, Schweinsburg A, Tapert S. 2009. “Altered white matter microstructure in adolescent substance users” Psychiatry Res 173(3): 228-237
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2734872/
36 marijuana and alcohol-using (MJ+ALC) adolescents (ages 16-19) and 36 demographically similar non-using controls evaluated with DTI
MJ+ALC teens had significantly lower FA than controls in 10 regions, including left superior longitudinal fasciculus (SLF), left postcentral gyrus, bilateral crus cerebri, and inferior frontal and temporal white matter tracts. These diminutions occurred in the context of increased FA in right occipital, internal capsule, and SLF regions. Changes in MD were less distributed, but increased MD was evident in the right occipital lobe, whereas the left inferior longitudinal fasciculus showed lower MD in MJ+ALC users
Studies examining brain morphology in marijuana users, particularly within white matter, offer equivocal findings. Although some report no changes in white matter volume and composition in adult users (Block et al., 2000; Gruber and Yurgelun-Todd, 2005), reduced white matter volumes in the left parietal lobe and increased tissue density surrounding the left parahippocampal and fusiform gyri have been documented (Matochik et al., 2005). Whether the reported changes in brain structure among marijuana using adolescents persist into adulthood remains tentative. Increased mean diffusivity in the prefrontal fiber bundles of the corpus callosum in adults who initiated use during early adolescence suggests long-term changes to white matter quality as a result of adolescent marijuana use (Arnone et al., 2008). In conflict with these findings, a DTI study using whole-brain voxelwise analysis of 10 young adults who used moderately as adolescents suggested no loss of white matter integrity relative to non-users (Delisi et al., 2006). Still, another report shows higher fractional anisotropy in the rostral body and isthmus of the corpus callosum in adolescent substance users compared to controls (De Bellis et al., 2008)
Although cannabis can prevent oligodendrocyte death (Molina-Holgado et al., 2002b), early or chronic cannabis exposure may cause a down-regulation of CB-1 receptors and suppress oligodendrocyte function during neurodevelopment
pre-onset use white matter integrity in MJ+ALC users is unknown, and pre-existing vulnerabilities including increased incidence of behavioral dysregulation (Kirisci et al., 2004; Tarter et al., 2003) and negative affectivity (Chassin et al., 2004) may be associated with neurological differences that predispose individuals in this group to substance use. Further, MJ+ALC users were more likely to have a parental history of substance use disorders. Considering that white matter changes in adolescent MJ+ALC users may have predated the onset of use, future follow-ups of this cohort will help determine whether increases in substance use result in greater deviations in white matter microstructure
Beaulieu J-M, Sotnikova T, Marion S, Lefkowitz R, Gainetdinov R, Caron M. 2005. “An Akt/β-Arrestin 2/PP2A signaling complex mediates dopaminergic neurotransmission and behavior“ Cell 122(2): 261-273
http://www.sciencedirect.com/science/article/pii/S0092867405004575
β-arrestin 2 deficiency in mice results in reduction of dopamine-dependent behaviors, loss of Akt regulation by dopamine in the striatum, and disruption of the dopamine-dependent interaction of Akt with its negative regulator, protein phosphatase 2A. Importantly, canonical cAMP-mediated dopamine-receptor signaling is not inhibited in the absence of β-arrestin 2. These results demonstrate that, apart from its classical function in receptor desensitization, β-arrestin 2 also acts as a signaling intermediate through a kinase/phosphatase scaffold
Dysregulation of dopaminergic neurotransmission is associated with multiple neurological and psychiatric conditions such as Parkinson’s disease, Huntington’s disease, attention deficit hyperactivity disorder (ADHD), mood disorders, and schizophrenia (Carlsson, 2001 and Gainetdinov and Caron, 2003). In the mammalian brain, most dopaminergic neurons project from the substantia nigra to the striatum, a major area of basal ganglia mostly populated by medium spiny neurons that are responsive to DA (Greengard, 2001). Two classes of GPCR have been shown to mediate all DA functions. D1 class receptors (D1R and D5R) are mostly coupled to Gαs and positively regulate the production of the second messenger cAMP and the activity of its main neuronal target, protein kinase A (PKA) (Missale et al., 1998). D2 class receptors (D2R, D3R, and D4R) couple to Gαi/o, thus downregulating cAMP production and PKA activity (Missale et al., 1998). Moreover, this class of DA receptors can also modulate intracellular Ca2+ levels (Missale et al., 1998 and Nishi et al., 1997), leading to changes in the activity of Ca2+-regulated signaling proteins such as the protein phosphatase calcineurin (PP2B) (Nishi et al., 1997)
prolonged stimulation of D2 class receptors leads to specific dephosphorylation/inactivation of the serine/threonine kinase Akt on its regulatory Thr308 residue (Beaulieu et al., 2004). Interestingly, phosphorylation of Akt on its second regulatory residue (Ser473) was not affected by DA (Beaulieu et al., 2004). Regulation of Akt by DA was observed in the striatum of mice that display persistently elevated levels of extracellular DA due to a lack of the DA transporter (DAT-KO mice). Similar changes in Akt phosphorylation also occurred in normal mice (wt) treated with direct or indirect DA agonists such as apomorphine or amphetamine (Beaulieu et al., 2004). Moreover, depletion of striatal DA in DAT-KO mice or D2 class-receptor blockade resulted in enhanced phosphorylation/activation of Akt, thus indicating that D2 class receptors are responsible for the regulation of Akt by DA (Beaulieu et al., 2004 and Emamian et al., 2004)
Dephosphorylation of Akt in response to DA leads to a reduction of kinase activity and a concomitant activation of its substrates glycogen synthase kinase 3 α and β (GSK3α and GSK3β) since both are negatively regulated by Akt (Beaulieu et al., 2004 and Cross et al., 1995)… mice lacking one allele of the GSK3β gene show markedly reduced locomotor responses to amphetamine (Beaulieu et al., 2004), while mice lacking the Akt isoform Akt1 display behavioral manifestations generally associated with enhanced dopaminergic responses (Emamian et al., 2004),
GPCR activation and their phosphorylation by GPCR kinases (GRKs), β-arrestins bind to the receptors to uncouple them from G proteins and participate in the recruitment of the endocytic protein complex, thus leading to an attenuation of GPCR signaling (Gainetdinov et al., 2004)
most behavioral actions of DA are thought to be associated with the modulation of adenylate cyclase and PKA activity (Greengard, 2001 and Missale et al., 1998)
we have shown that a β-arrestin 2-mediated kinase/phosphatase scaffolding of Akt and PP2A is responsible for the regulation of Akt by DA receptors
Beaulieu J-M, Gainetdinov R, Caron M. 2007. ‘The Akt-GSK-3 signaling cascade in the actions of dopamine” Trends Pharmacol Sci 28(4): 166-72
http://www.ncbi.nlm.nih.gov/pubmed/17349698
Classically, dopamine receptors have been shown to regulate cAMP-PKA (protein kinase A) and Ca(2+) pathways through G-protein-mediated signaling. However, it has become apparent that, in addition to this canonical action, D(2)-class dopamine receptors can function through a protein kinase B (Akt)-GSK-3 (glycogen synthase kinase 3) signaling cascade
Bechtold J, Simpson T, White H, Pardini D. 2015. “Chronic adolescent marijuana use as a risk factor for physical and mental health problems in young adult men” Psychology of Addictive Behaviors 29(3): 552-563
http://www.apa.org/pubs/journals/releases/adb-adb0000103.pdf
Findings from latent class growth curve analysis identified 4 distinct subgroups of marijuana users: early onset chronic users, late increasing users, adolescence-limited users, and low/nonusers. Results indicated that the 4 marijuana use trajectory groups were not significantly different in terms of their physical and mental health problems assessed in the mid-30s
Becker M, Collins P, Luciana M. 2014. “Neurocognition in college-aged daily marijuana users” Journal of Clinical and Experimental Neuropsychology 36(4): 379-398
http://www.ncbi.nlm.nih.gov/pubmed/24620756
Marijuana users were high functioning, demonstrating comparable IQs to controls and relatively better processing speed. Marijuana users demonstrated relative cognitive impairments in verbal memory, spatial working memory, spatial planning, and motivated decision making. Comorbid use of alcohol, which was heavier in marijuana users, was unexpectedly found to be associated with better performance in some of these areas
Behan B, Connolly C, Datwani S, Doucet M, Ivanovic J, Morioka R, Stone A, Watts R, Smyth B, Garavan H. 2013. “Response inhibition and elevated parietal-cerebellar correlations in chronic adolescent cannabis users” Neuropharmacology 84:131-7
http://www.ncbi.nlm.nih.gov/pubmed/23791961
a cohort of adolescent heavy cannabis users and age-matched non-cannabis -using controls completed a Go/No-Go paradigm. Users were impaired in performance on the task but voxel-wise and region-of-interest comparisons revealed no activation differences between groups. Instead, an analysis of correlation patterns between task-activated areas revea led heightened correlation scores in the users between bilateral inferior parietal lobules and the left cerebellum. The increased correlation activity between these regions was replicated with resting state fMRI data
Seventeen current cannabis users (1 female, mean age: 16.5, range 15 e18) and 18 comparison subjects (1 female, mean age: 16.1, range 14 e19) took part in the experiment. Participants were screened for no history of neurological/psychiatric illness or any past loss of consciousness which required hospitalization. All partic-ipants were right-handed. Users were recruited from several drug treatment centres in Dublin, Ireland and were being treated for cannabis dependency at the time of scanning
Groups significantly differed (t(33) ¼ 2.938, p < 0.01) on intellectual ability (as measured by the word reading section from the wide range achievement test 4 (WRAT 4)) with cannabis users displaying lower scores. Cannabis users also exhibited higher scores for current levels of stress (t(32) ¼ 2.20 4, p < 0.05) and anxiety (t(32) ¼ 2.228, p < 0.05) as indexed by the Depression Anxiety Stress Scales (DASS)… the cannabis-using group contained more smokers and these had smoked more cigarettes in the week and month prior to participation
increased correlations between parietal and cerebellar regions. This effect was replicated in resting state fMRI data acquired during the same session indicating that the significant correlation pattern between the parietal and cerebellar regions was not solely a response to a particular task demand but may, instead, re fl ect an intrinsic feature of the connectivity of the adolescent cannabis user ’s brain
A potentially unfavourable over-reliance on the cerebellum during cognitive task s has been previously demonstrated in cocaine users ( Hester and Garavan, 20 0 4)… Desmond et al., 20 03 reported that alcohol-dependent subjects demonstrated increased activation of the right cerebellum, as well as the lef t prefrontal cortex, during a working memory task and maintained similar performance levels to controls… the cerebellum might compensate when other task-related regions are not engaged… we identified lower correlation scores within the cannabis-using group between networks comprising right inferior frontal and right middle frontal gyri… the adolescent cannabis users in the present study did not show increased activity in the cerebellum, only increased correlations with fronto-parietal cortex… with CB1 receptors highly expressed in the cerebellum and previous reports indicating that cerebellar-mediated processes such as eyeblink conditioning are disrupted in young adult cannabis users, any pu-tative compensatory mechanism involving the cerebellum may be disrupted by heavy cannabis use (Quickfall and Crockford, 20 0 6; Skosnik et al., 20 08 ).
cerebral blood volume measures, taken at rest, have been recorded in adult cannabis users following cessation of use (Sneider et al., 20 0 6, 20 08). At day 0 and following seven days of abstinence, users exhibited increased blood volume differences in frontal, temporal and cerebellar areas. However, following 28 days of abstinence, only a lef t temporal area and the cerebellum still displayed increased blood volume rates in users. Such long-lasting cerebellar alterations have been supported by previous structural imaging work demonstrating increased grey matter vermis volumes in adolescent cannabis users following one month of abstinence (Medina et al., 2010 ). Similar differences in cerebellar structure in adult users have also been noted ( Cousijn et al., 2012). It also appears that development of the cerebellum is not as heavily determined by genes as are other brain regions and thus this structure may be particularly vulnerable to environmental conditions (Wallace et al., 20 0 6).
it is acknowledged that the use of other drugs may have impacted upon response inhibition functioning. For example, two recent studies have demonstrated that adult smokers displayed signifi cantly poorer performance levels on Go/No-Go task s (Luijten et al., 2011; Nestor et al., 2011). … there may be some argument that these are not adequate sample sizes for such an investigation
Beltramo M, de Fonseca F, Navarro M, Calignano A, Gorriti M, Grammatikopoulos G, Sadile A, Giuffrida A, Piomelli D. 2000. “Reversal of dopamine D(2) receptor responses by an anandamide transport inhibitor” J Neurosci 20(9): 3401-7
http://www.ncbi.nlm.nih.gov/pubmed/10777802
We characterized the pharmacological properties of the anandamide transport inhibitor N-(4-hydroxyphenyl)-arachidonamide (AM404) in rats and investigated the effects of this drug on behavioral responses associated with activation of dopamine D(2) family receptors
AM404 caused a mild and slow-developing hypokinesia that was significant 60 min after intracerebroventricular injection of the drug and was reversed by the CB1 cannabinoid receptor antagonist SR141716A
AM404 prevented the stereotypic yawning produced by systemic administration of a low dose of apomorphine, an effect that was dose-dependent and blocked by SR141716A. Furthermore, AM404 reduced the stimulation of motor behaviors elicited by the selective D(2) family receptor agonist quinpirole. Finally, AM404 reduced hyperactivity in juvenile spontaneously hypertensive rats, a putative model of attention deficit hyperactivity disorder. The results support a primary role of the endocannabinoid system in the regulation of psychomotor activity and point to anandamide transport as a potential target for neuropsychiatric medicines
Bénard G, Massa F, Puente N, Lourenco J, Bellocchio L, Soria-Gomez E, Matias I, Delamarre A, Metna-Laurent M, Cannich A, Hebert-Chatelain E, Mulle C, Ortega-Gutierrez S, Martin-Fontecha M, Klugmann M, Guggenhuber S, Lutz B, Gertsch J, Chaouloff F, Lopez-Rodriguez M, Grandes P, Rossignol R, Marsicano G. 2012. “Mitochondrial CB1 receptors regulate neuronal energy metabolism” Nat Neurosci 15(4): 558-64
http://www.ncbi.nlm.nih.gov/pubmed/22388959 add to dropbox
The mammalian brain is one of the organs with the highest energy demands, and mitochondria are key determinants of its functions. Here we show that the type-1 cannabinoid receptor (CB1) is present at the membranes of mouse neuronal mitochondria (mtCB1), where it directly controls cellular respiration and energy production. Through activation of mtCB1 receptors, exogenous cannabinoids and in situ endocannabinoids decreased cyclic AMP concentration, protein kinase A activity, complex I enzymatic activity and respiration in neuronal mitochondria. In addition, intracellular CB1 receptors and mitochondrial mechanisms contributed to endocannabinoid-dependent depolarization-induced suppression of inhibition in the hippocampus
The brain represents only 2% of the total body weight in mammals, but it consumes up to 20% of the body’s resting energy production
several reports have shown the intramitochondrial localization of potential downstream effectors of G protein signaling, such as soluble adenylyl cyclase, phosphodiesterase, and protein kinase A (PKA). Thus, cAMP can be produced in mitochondria, leading to activation of PKA and phosphorylation of mitochondrial proteins, eventually regulating mitochondrial respiration and energy production… mechanisms coupling mitochondrial activity and neuronal physiology, remain poorly understood
CB1 receptor signaling typically involves reduction of cAMP, and mitochondrial forms of adenylyl cyclase and protein kinase A (PKA) have been suggested to regulated complex I activity and mitochondrial respiration. Acute THC treatment decreased mitochondrial cAMP by approximately 30%
Brain mitochondria have substantial levels of endocannabinoid degrading activity
only the activation of intracellular CB1 receptors affects mitochondrial respiration of living cells
CB1 GPCR is present in a portion (approximately 30% in the hippocampus) of brain neuronal mitochondria
The open source program MitoProt II (ref 35) predicted the CB1 receptor protein to bear a 39.3% probability of mitochondrial localization. This theoretical value is much higher than that of many GPCRs and is comparable to the values of several known or recently identified mitochondrial proteins. … The proportion of CB1 receptors located on the mitochondrial membrane (approximately 15% of total cell CB1 receptors) might seem low. However, this expression accounts for up to 30% of mtCB1 dependent reduction of respiration in purified brain mitochondria.
Although a minimal amount (less than 10% of total cortical CB1 receptor proteins are expressed in glutamatergic neurons, many of the functions of the endocannabinoid system are exerted by this limited percentage of receptors. For instance, protection against excitotoxic seizures or the orexigenic properties of endocannabinoid signaling are mainly exerted by glutamatergic-neuron CB1 receptors
DSI is short-lasting (a few tens of seconds), likely due to efficient endogenous degradation of endocannabinoids. Conversely, drug applications (especially of lipphilic cannabinoids) are generally long-lasting, their duration depending only on pharmacokinetics properties of the drug, and might induce extra effects. From the spatial point of view, the moilization of endocannabinoids by depolarization and/or synaptic stimulation might be limited to specific cellular subdomains. Conversely, exogenous cannabinoids might display preference for other subdomains, thereby triggering different mechanisms. Finally, depolarization steps able to endogenously mobilize endocannabinoids might trigger other, so far unknown cellular events to favor additional means of endocannabinoid signaling, which would otherwise remain silent.
Berk M, Williams L, Jacka F, O’Neil A, Pasco J, Moylan S, Allen N, Stuart A, Hayley A, Byrne M, Maes M. 2013. “So depression is an inflammatory disease, but where does the inflammation come from?” BMC Medicine 11:200
http://www.biomedcentral.com/1741-7015/11/200
A range of factors appear to increase the risk for the development of depression, and seem to be associated with systemic inflammation; these include psychosocial stressors, poor diet, physical inactivity, obesity, smoking, altered gut permeability, atopy, dental cares, sleep and vitamin D deficiency
the above mentioned sources of inflammation may play a role in other psychiatric disorders, such as bipolar disorder, schizophrenia, autism and post-traumatic stress disorder
There is now an extensive body of data showing that depression is associated with both a chronic low-grade inflammatory response, activation of cell-mediated immunity and activation of the compensatory anti-inflammatory reflex system (CIRS), characterized by negative immunoregulatory processes [1,2]. New evidence shows that clinical depression is accompanied by increased oxidative and nitrosative stress (O&NS) and autoimmune responses directed against O&NS modified neoepitopes [3,4].
cytokines induce depressive-like behaviors; in studies where healthy participants are given endotoxin infusions to trigger cytokines release, classical depressive symptoms emerge [7]. Exogenous cytokine infusions also cause the classical phenotypic behavioral and cognitive features of depression. As an exemplar, a quarter of the people given interferon for the treatment of hepatitis C develop emergent major depression [8,9]
remission of clinical depression is accompanied by a normalization of inflammatory markers [15], while lack of response is associated with persistently elevated levels of inflammatory markers [16]
These data beg the question
many inflammatory disorders (chronic obstructive pulmonary disease, cardiovascular disease (CVD) and autoimmune disorders) and neuroinflammatory disorders (multiple sclerosis and Parkinson’s disorder) and inflammatory conditions (hemodialysis and the postpartum period) may trigger clinical depression [17]. However, these factors are only present in a small percentage of the larger population of depressed individuals
Psychosocial stressors, including acute psychological trauma or more sub-chronic stressors, and early exposure to childhood trauma robustly increase the risk of developing clinical depression and mood symptoms, while impacting neuro-immune circuits
In humans, there is evidence that different types of psychosocial stressors may stimulate the pro-inflammatory cytokine network, including increases in IL-6 and TNFα [25–28]. Maes et al. [28,29] were the first to report that stress-induced increases in IFNγ and stress-induced Th1 dominance were significantly correlated with stress-induced anxiety and distress. Thus, subjects with psychological stress-induced distress and anxiety showed significantly greater increases in IFNγ and lower IL-10 than those without distress and anxiety. Psychosocial stress is also accompanied by lowered levels of endogenous, anti-inflammatory compounds, for example, CC16 (uteroglobuline), which decreases the production of IFNγ [30]. Individuals showing stress-induced decreases in CC16 in the serum display higher stress-induced anxiety and distress, and an increased production of IFNγ during the stress condition [29,30]. Thus, stress-induced increases in pro-inflammatory and Th1-like cytokines may be mediated by lowered levels of endogenous anti-inflammatory compounds, such as CC16. Stress-induced production of pro-inflammatory cytokines, for example, TNFα and IL-6, and Th1-like cytokines, for example, IFNγ, are related to an increased number of leukocytes and neutrophils, and expression of immune cell activation markers, including CD2+CD26+ and CD2+HLADR, and different signs of an acute phase response [29]. This indicates that psychosocial stress-induced elevations in pro-inflammatory cytokines orchestrate stress-induced changes in peripheral blood immune cells, inflammatory reactions and neurobehavioral changes
psychosocial stressors and acute psychotrauma may trigger mood disorders in vulnerable subjects, for example, those with immune gene polymorphisms, lowered levels of pepdidases, including dipeptidylpeptidase and prolylendopeptidase, and those with increased inflammatory burden [31]
individuals experiencing stress in childhood resulting from maltreatment, abuse, social isolation and economic hardship are twice as likely to suffer chronic inflammation [32]. The detrimental impact of adversity on health in adulthood has also been demonstrated in US populations. Kiecolt-Glaser [33] found that childhood adversity can shorten the lifespan by 7 to 15 years, arguing that stress associated with abuse, death of a parent or parental relationship problems can lead to inflammation and premature cell aging, when compared with individuals who have not experienced such adversity. Miller et al. [34], in a further study focusing on depression outcomes, compared C-Reactive Protein (CRP) and IL-6 levels of women with and without history of childhood adversity; the former group was shown to have a greater likelihood of depression, recording higher levels of inflammation using these biomarkers. Studies exploring the influence of stress on other inflammatory diseases, such as CVD [35] and metabolic syndrome [36], have consistently shown similar trends
the association between childhood adversity and vulnerability to inflammatory disease cannot fully be explained by a prolonged period of stress initiated by such an event. Rather, it is possible that learned, maladaptive responses to stress occurring in early childhood are also employed later in adult life in response to stressors. Thus, stress in adulthood has become of increasing interest as an instrumental risk factor for disease onset
Slavich et al. [38] found that responses to social stress via neural activity lead to marked increases in inflammatory activity. Similarly, Emeny[39] found job strain to have a direct effect on inflammation, and to influence other risk factors for inflammation. Job strain is known as a risk factor for other inflammatory diseases, such as CVD, and more recently has been shown to be strongly associated with depression risk [40]. Indeed, it is clear that understanding modifiable risk factors related to stress (and lifestyle) may be an important step in the prevention of inflammatory diseases like depression
habitual or regular exercise protects against the development of new depressive illnesses [82–84], and that physical inactivity during childhood is associated with an increased risk of depression in adulthood [85]
Acute exercise generates reactive oxygen species (ROS) [93] and inflammatory cytokines [94] that can transiently damage muscle cells, causing muscle fatigue, pain and inflammation. Contracting skeletal muscle produces a number of ‘myokines’, such as IL-6 [95], which impact systemically on lipid and glucose metabolism [96]. The pattern of inflammatory markers produced during acute exercise, characterized by a rapid elevation in levels of IL-6 that is quickly followed by induction of anti-inflammatory substances, including IL-1ra, IL-10 and soluble tumor necrosis factor receptor (sTNF-R) [97], differs markedly from that in other inflammatory conditions, such as sepsis. Recovery after the exercise-induced IL-6 spike dampens the inflammatory response and oxidative burst activity[98]. Chronic or regular exercise, therefore, down-regulates systemic inflammation via homeostatic adaptation [99]. Similarly, fitness and exercise reduces leptin [100], elevated levels of which are also implicated in the development of depression [101] and is the most evidence-based management strategy for insulin resistance [102]
Depression has also been shown to predispose to obesity in a bidirectional manner [112]. A recent meta-analysis of prospective cohort studies found obesity to increase the risk of later depression by 55%, while depression increased the risk of developing obesity by 58% [113]
Altered adipocyte function, fatty acid levels, leptin and hypothalamic pituitary adrenal (HPA) axis dysfunction and oxidative stress are hypothesized to play a crucial but synergistic role in obesity-associated inflammation [114]
Clinical depression has recently been shown to be accompanied by increased plasma levels of immunoglobulin (Ig) A and/or IgM directed against a number of gram negative bacteria, including Hafnia alvei, Pseudomonas aeruginosa,Morganella morganii, Proteus mirabilis, Pseudomonas putida, Citrobacter koseri and Klebsielle pneumoniae[138–140]. All these gram negative bacteria belong to the normal gut flora [141,142]
An increased permeability of the gut wall may allow poorly invasive gram negative bacteria to translocate into the mesenteric lymph nodes (MLNs) and sometimes into the systemic circulation [148,149]. Consequently, in the systemic circulation, IgM and IgA responses are mounted against the LPS of the bacterial wall, while IgA responses may be mounted even when the bacteria do not reach the blood stream, but only translocate into the MLNs
IgE response to common allergen exposure, leading to the development of allergic symptoms, such as asthma, eczema or allergic rhinitis/hay fever is defined as atopy [157]. The prevalence of atopic disorders has been steadily increasing over the past few decades [158,159]. Interestingly, atopy and depression have recently been linked. Although methodologies differ among studies, it has been consistently reported that atopic disorders are associated with an increased risk of both clinical depression and depressive symptomatology in clinical settings [160–163]. Population-based studies provide further support, showing a positive association between depression and atopic disorders [164–168]
It is estimated that up to 80 to 90% of individuals who suffer from a MDD also experience sleep disturbances [194–196]
Smith and Maes in 1995 launched the monocyte-T lymphocyte theory of schizophrenia, which considered that activation of immuno-inflammatory processes may explain the neurodevelopmental pathology related to gestational infections. Results of recent meta-analyses showed that schizophrenia is accompanied by activation of inflammatory and cell mediated pathways [234]. PTSD patients also show higher levels of pro-inflammatory cytokines, including IL-1[235], IL-6 [236,237] and TNFα [238]
he strong associations among schizophrenia and smoking [240], obesity [241], some atopic disorders [242], sleep disorders [243] and poor periodontal and oral health [244,245] may further contribute to the inflammatory burden in schizophrenia patients
Berman R, Cappiello A, Anand A, Oren D, Heninger G, Charney D, Krystal J. 1999. “Antidepressant effects of ketamine in depressed patients” Biological Psychiatry 47(4): 351-354
http://www.biologicalpsychiatryjournal.com/article/S0006-3223(99)00230-9/abstract
Bersani G, Orlandi V, Kotzalidis G, Pancheri P. 2002. “Cannabis and schizophrenia: impact on onset, course, psychopathology and outcomes” Eur Arch Psychiatry Clin Neurosci 252(2): 86-92
http://www.ncbi.nlm.nih.gov/pubmed/12111342
Family history positive for psychosis was more frequent in consumers, especially when consumption started before onset of schizophrenia. Subjects whose onset of schizophrenia preceded the beginning of cannabis abuse had more positive symptoms than those who started abuse before the onset of schizophrenia
had less negative symptoms than nonabusers
Beylin A, Shors T. 2003. “Glucocorticoids are necessary for enhancing the acquisition of associative memories after acute stressful experience” Horm Behav 43(1): 124-131
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3363955/
Exposure to acute stressful experience can enhance the later ability to acquire new memories about associations between stimuli. This enhanced learning is observed during classical eyeblink conditioning of both hippocampal-dependent and -independent learning. It can be induced within minutes of the stressful event and persists for days. Here we examined the role of the major stress hormones glucocorticoids in the enhancement of learning after stress
one injection of stress levels of corticosterone enhanced new learning within minutes but not new learning 24 h later
In the case of glucocorticoids, exogenous administration enhances some processes of learning (Sandi and Rose, 1997; Roozendaal and McGaugh, 1997b), whereas removal via adrenalectomy (ADX) has been associated with learning impairments (Conrad and Roy, 1995;Pugh et al., 1997; Vaher et al., 1994). In previous studies, we found a positive correlation between basal levels of glucocorticoids during training and acquisition of trace memories in male rats (Wood et al., 2001). The medullary substances epinephrine and norepinephrine have also been associated with enhanced performance on learning tasks, whereas antagonism of their receptors can be detrimental (Gamaro et al., 1997; Leon, 1998; Roozendaal et al., 1996)
stress-induced enhancement of conditioning can be mimicked by an acute exposure to glucocorticoids, but the effect does not persist, as it does after exposure to the acute tailshock stress
training with both delay and trace conditioning increases the observation of dendritic spines in area CA1 of the hippocampus (Leuner et al., 2002)
reports that glucocorticoids impair processes of retrieval and in some cases acquisition, at least during spatial navigation learning (de Quervain et al., 1998;Diamond and Rose, 1994; Kim et al., 2001; Kim and Diamond, 2002)
Bhattacharyya S, Atakan Z, Martin-Santos R, Crippa J, Kambeitz J, Prata D, Williams S, Brammer M, Collier D, McGuire P. 2012. “Preliminary report of biological basis of sensitivity to the effects of cannabis on psychosis: AKT1 and DAT1 genotype modulates the effects of d-9-tetrahydrocannabinol on midbrain and striatal function” Molecular Psychiatry 17: 1152-1155
http://www.ncbi.nlm.nih.gov/pubmed/22290123
Cannabis use can induce acute psychotic symptoms 1 and increase the long-term risk of schizophrenia2
The human DAT1 gene has a polymorphic 40 bp variable number of tandem repeats (VNTR) in the 3′ untranslated region (UTR), with several alleles ranging from 3 to 11 copies of the 40-bp repeats, the 9-repeat and 10-repeat alleles being the most common. The DAT1 3′ UTR VNTR modulates activity in the striatum 7,9 and midbrain, 9 and its 9-repeat allele is asso-ciated with higher synaptic dopamine levels than the 10-repeat counterpart. 10 AKT1 has recently been impli-cated in mediating the effects of cannabis on psycho-sis, 11 and variants of both genes have been linked with schizophrenia in genetic association studies.12–14
35 healthy volunteers who had minimal previous cannabis use
There was a main effect of DAT1 genotype (F1,31 = 7.46, P = 0.01; 9-repeat carriers > 10-repeat homo-zygotes) and a trend for a main effect of AKT1 genotype (F 1,31 =3.42, P = 0.07; G homozygotes > heterozygotes) on the increase in psychotic symptoms induced by d-9-THC. There was also an interaction (F1,31 =4.49, P = 0.04) between the effects of the DAT1 and AKT1 genotypes on the symptomatic effect of d-9-THC; the increase in symptoms was greater in individuals who were G homozygotes of AKT1 rs1130233 and also carriers of the 9-repeat allele of the DAT1 3′ UTR VNTR (hereafter referred to as GG/9-repeat carrier; n =7), relative to those that were not
During encoding, following placebo, indi-viduals who were G homozygotes of AKT1 and carriers of the 9-repeat allele (DAT1 3 ‘ UTR VNTR) showed strong striatal activation, whereas there was a little response in the REST. d9 THC markedly attenuated the striatal activation in GG/9-repeat carrier individuals, but had a minimal effect in the REST group. Similarly, during the recall condition, following placebo, subjects who were homozygotes for the G allele of AKT1 and carriers of the 9-repeat allele for the DAT1 3’UTR VNTR showed strong activation in the midbrain, whereas there was only a modest response in the REST. Administration of d9-THC led to deactivation in the midbrain in GG/9-repeat carrier individuals, but had little effect on the response in the REST group
there were no significant interactions between d9-THC and genotype on task performance
the more d9-THC attenuated striatal and midbrain activation, the greater the severity of the psychotic symptoms
Blaauw E, Arensman E, Kraaij V, Winkel F, Bout R. 2002. “Traumaatic life events and suicide risk among jail inmates: the influence of types of events, time period and significant others” J Trauma Stress 15(1): 9-16
http://www.ncbi.nlm.nih.gov/pubmed/11936726/
Relationships between traumatic life events and suicide risk were studied in two samples of jail inmates with a low (N = 216) and a high (N = 51) suicide risk. Although nonsuicidal inmates reported a high prevalence of traumatic life events, suicidal inmates reported even higher prevalence rates. Suicidal inmates reported more episodes of sexual abuse, physical maltreatment, emotional maltreatment, abandonment, and suicide attempts by significant others. They also had experienced more traumatic life events during childhood, later life, and detention
Black, C. 2013. The Flight of the Eagle: A Strategic History of the United States. Signal.
http://books.google.ca/books/about/Flight_of_the_Eagle.html?id=9c7rTGbHj-QC&redir_esc=y
Describing the United States in 2013: “its justice system was corrupt and oppressive and produced 6 to 12 times the per capita number of incarcerated people as other prosperous democracies, on behalf of a war on drugs that is an exercise in hypocrisy and futility in which America has been more decisively defeated than in any war it actually waged. A rogue prosecutocracy terrorizes the country; it wins 99.5 per cent of its cases, 97 percent without a trial, so stacked is the judicial deck and withered the guarantees of individual liberties in the Bill of Rights. Forty-eight million Americans have a criminal record and there is minimal general recognition of the evils of this system.”
Blair R. 2001. “Neurocognitive models of aggression, the antisocial personality disorders, and psychopathy” J Neurol Neurosurg Psychiatry 71: 727-731
http://jnnp.bmj.com/content/71/6/727.full
Impairments in executive emotional systems (the somatic marker system or the social response reversal system) are related to reactive aggression shown by patients with “acquired sociopathy” due to orbitofrontal cortex lesions
“Acquired sociopathy” was a term introduced to characterise persons who, after acquired lesions of the orbitofrontal cortex, fulfil the DSM-III diagnostic criteria for “sociopathic disorder”, an antecedent of APD.2
Individuals with acquired sociopathy following orbitofrontal cortex lesions are a relatively homogeneous population; they present solely with reactive aggression even if the lesion occurs early in life
it is damage to the orbitofrontal cortex, rather than any other region of the frontal or posterior cortex, which places the patient at greatest risk of presenting with reactive aggression.17
The instrumental antisocial behaviours shown by the developmental psychopath are a consequence of inability to socialise due to an impairment in the capacity to form associations between emotional unconditioned stimuli (particularly distress cues) and conditioned stimuli (specifically representations of transgressions). If the person is raised in a social environment (for example, poverty) where there are advantages for engaging in antisocial behaviour, they may engage in this behaviour but will not experience aversion to the distress of their victims. Because of the heterogeneity in the populations of those with CD and APD, different cases are likely to correspond to either a developmental form of acquired sociopathy (and some cases may reflect trauma) or developmental psychopathy itself
Blanco C, Hasin D, Wall M, Florez-Salamanca F, Hoertel N, Wang S, Kerridge B, Olfson M. 2016. “Cannabis use and risk of psychiatric disorders Prospective evidence from a US National Longitudinal Study” JAMA Psychiatry doi:10.1001/jamapsychiatry.2015.3111
http://archpsyc.jamanetwork.com/article.aspx?articleid=2491944
We used multiple regression and propensity score matching to estimate the strength of independent associations between cannabis use at wave 1 and incident and prevalent psychiatric disorders at wave 2. Psychiatric disorders were measured with a structured interview (Alcohol Use Disorder and Associated Disabilities Interview Schedule–DSM-IV). In both analyses, the same set of wave 1 confounders was used, including sociodemographic characteristics, family history of substance use disorder, disturbed family environment, childhood parental loss, low self-esteem, social deviance, education, recent trauma, past and present psychiatric disorders, and respondent’s history of divorce
34 653 respondents (14 564 male [47.9% weighted]; mean [SD] age, 45.1 [17.3] years), cannabis use in wave 1 (2001-2002), which was reported by 1279 respondents, was significantly associated with substance use disorders in wave 2 (2004-2005) (any substance use disorder: odds ratio [OR], 6.2; 95% CI, 4.1-9.4; any alcohol use disorder: OR, 2.7; 95% CI, 1.9-3.8; any cannabis use disorder: OR, 9.5; 95% CI, 6.4-14.1; any other drug use disorder: OR, 2.6; 95% CI, 1.6-4.4; and nicotine dependence: OR, 1.7; 95% CI, 1.2-2.4), but not any mood disorder (OR, 1.1; 95% CI, 0.8-1.4) or anxiety disorder (OR, 0.9; 95% CI, 0.7-1.1)
Although the cross-sectional association between cannabis use and psychiatric disorders has been consistently documented,12,13 longitudinal studies evaluating the association of cannabis use with incident psychiatric disorders have yielded mixed results.14– 16 Some studies have reported that cannabis use is associated with increased risk of onset of depression,16– 19 anxiety,14,17,19 bipolar disorder,16 substance use disorders,20 and psychosis,21,22 while others have not replicated these findings.14– 16,23 Discrepancies may be partly explained by a focus on varying age ranges,14,15,17– 19,23– 25 geographic locations,18,23– 25 males vs females,15 or number and type of mental disorders evaluated.14– 19,23– 26
Confounders from the childhood tier included family history of SUDs (lifetime history of alcohol or drug use disorders in the biological parents or siblings), parental loss (parent’s divorce or death of at least 1 parent before the participant was 18 years old), and vulnerable family environment (parental absence or separation from a biological parent before age 18 years). The confounders for the early-adolescence tier included low self-esteem (dichotomous; scored 1 if respondents believed they were not as good, smart, or attractive as most other people), age at onset of anxiety disorders (with childhood onset before age 18 years), and social deviance (assessed as the number of conduct disorder or antisocial personality disorder behaviors in which the respondent engaged before age 15 years; range, 0-33). Confounder in the late-adolescence tier included educational level (in years), number of personality disorders, and number of Axis I disorders with onset before age 18 years. Confounders in the adulthood tier included history of divorce, history of SUD (alcohol use disorder, nicotine dependence, and drug use disorder other than cannabis use disorders), and social deviance (measured as the number of antisocial personality disorder behaviors in which the individual engaged after age 15 years but prior to the wave 1 assessment)
In the full sample of 34 653 respondents (14 564 male [47.9% weighted]; mean [SD] age, 45.1 [17.3] years), the 1279 individuals with cannabis use (772 male [66.4% weighted]; mean [SD] age, 29.9 [10.7] years) had greater odds of most risk predictors examined except age and history of divorce, which were lower among individuals with cannabis use, and educational level and nonwhite race/ethnicity, which did not differ across groups (eTable 1 in the Supplement). Among the individuals with cannabis use, 345 (28.4% weighted) had cannabis abuse in the past year and 99 (7.7%) had cannabis dependence in the past year
in unadjusted analyses in the full population, cannabis use was associated with increased prevalence and incidence of a broad range of psychiatric disorders. However, after adjusting for several covariates that predicted cannabis use, multiple regression analysis and propensity score matching converged in indicating that cannabis use was associated only with increased prevalence and incidence of alcohol and drug use disorders, including nicotine dependence
The new results also help reconcile discrepant findings from prior studies by suggesting that most of the association of cannabis use with non-SUD psychiatric disorders is explained by differences in the distribution of confounders between cannabis users and nonusers.14,15,17– 19,23– 25 In addition, our findings support a dose-response association between cannabis use and the risk of SUDs.16,17
our study does not establish a causal association between cannabis use and new onset of disorders because of the possibility of residual confounding, particularly confounders that may vary over time across survey waves.
Our study indicates that cannabis use is associated with increased prevalence and incidence of substance use disorders. These adverse psychiatric outcomes should be taken under careful consideration in clinical care and policy planning.
Blázquez C, Chiarlone A, Bellocchio L, Resel E, Pruunsild P, Garcia-Rincon D, Sentner M, Timmusk T, Lutz B, Galve-Roperh I, Guzman M. 2015. “The CB1 cannabinoid receptor signals striatal neuroprotection via a PI3K-Akt/mTORC1/BDNF pathway” Cell Death Differ 22(1): 1618-1629
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4563779/
CB1 receptor engagement protects striatal cells from excitotoxic death via the phosphatidylinositol 3-kinase/Akt/mammalian target of rapamycin complex 1 pathway, which, in turn, (2) induces brain-derived neurotrophic factor (BDNF) expression through the selective activation of BDNF gene promoter IV, an effect that is mediated by multiple transcription factors
protective effect of cannabinoids in those two experimental systems was prevented by the CB1-selective antagonist SR141716 (rimonabant) or upon blockade of the PI3K/Akt/mTORC1/BDNF pathway
we evaluated the effect of cannabinoids on the best characterized Bdnf gene promoters by using exon-specific qPCR primers. THC upregulated total BDNF transcripts (Ct=23) and, specifically, exon IV-containing BDNF transcripts
in striking contrast with the hippocampus and the striatum, BDNF expression in the mouse cortex, which expresses high levels of the CB1 receptor,1 was unaffected by either THC administration19, 51 or CB1 receptor genetic ablation,19, 52 while another study found only very marginal increases in BDNF levels in the medial prefrontal cortex and the frontoparietal cortex upon THC injection to rats.55 Hence, albeit for hitherto unknown molecular reasons, BDNF expression seems to be much more refractory to CB1 receptor activation in the cortex than in the striatum or hippocampus.
BDNF may be a bona fide marker not only of HD neurodegeneration17 but also of CB1 receptor-evoked neuroprotection
Block R, Wittenborn J. 1984. “Marijuana effects on semantic memory: verification of common and uncommon category members” Psychological Reports 55: 503-512
http://www.amsciepub.com/doi/abs/10.2466/pr0.1984.55.2.503
Marijuana did not alter the normal difference in reaction time between common and uncommon examples of categories, suggesting that effects of marijuana on associations do not derive directly from underlying, general alterations of semantic memory retrieval. Marijuana’s effects were not influenced by the demands on memory retrieval or by providing advance information relevant to the required decisions, suggesting memory retrieval was not impaired by this dose of marijuana.
Block R, O’Leary D, Ehrhardt J, Augustinack J, Ghoneim M, Arndt S, Hall J. 2000. “Effects of frequent marijuana use on brain tissue and composition” Neuroreport 11(3): 491-6
http://www.ncbi.nlm.nih.gov/pubmed/10718301
in 18 current, frequent, young adult marijuana users and 13 comparable, non-using controls… no differences in grey or white matter. Lower ventricular CSF in MJ users
Bloomfield M, Morgan C, Egerton A, Kapur S, Curran H, Howes O. 2014. “Dopaminergic function in cannabis users and its relationship to cannabis-induced psychotic symptoms” Biol Psychiatry 75(6): 470-8
http://www.ncbi.nlm.nih.gov/pubmed/23820822
These findings indicate that chronic cannabis use is associated with reduced dopamine synthesis capacity and question the hypothesis that cannabis increases the risk of psychotic disorders by inducing the same dopaminergic alterations seen in schizophrenia
Bola J, Lehtinen K, Cullberg J, Ciompi L. 2009. “Psychosocial treatment, antipsychotic postponement, and low-dose medication strategies in first-episode psychosis: a review of the literature” Psychosis 1(1): 4-18
http://psychrights.org/research/Digest/Effective/PsychoSocialMoreEffective2009Psychosis.pdf
Five studies were ineludcd (N=261). each reporting modestly better long-term outcomes than initial medication treatment. resulting in a composite smallmedium effect-size advantage (r .= 0.17). In addition, 27-43% of experimental patients were not receiving anti psychotics at the two- or three-year follow-up
Bola J, Mosher L. 2003. “Treatment of acute psychosis without neuroleptics: two-year outcomes from the Soteria Project” Nervous and Mental Disease 191(4): 219-229
http://psychrights.org/research/Digest/Effective/bolamosher03.pdf
Newly diagnosed DSM-II schizophrenia subjects were assigned consecutively (1971 to 1976, N 79) or randomly (1976 to 1979, N 100) to the hospital or Soteria and followed for 2 years
Completing subjects with schizophrenia exhibited a large effect size benefit with Soteria treatment (.81 SD, p .02), particularly in domains of psychopathology, work, and social functioning. Soteria treatment resulted in better 2-year outcomes for patients with newly diagnosed schizophrenia spectrum psychoses, particularly for completing subjects and for those with schizophrenia. In addition, only 58% of Soteria subjects received antipsychotic medications during the follow-up period, and only 19% were continuously maintained on antipsychotic medications.
Bolla K, Eldreth D, Matochik J, Cadet J. 2005. “Neural substrates of faulty decision-making in abstinent marijuana users.” Neuroimage 26(2):480-92
http://www.ncbi.nlm.nih.gov/pubmed/15907305 in dropbox
A dose-related association was found between increased MJ use and lower IGT performance and alterations in brain activity. The MJ group showed greater activation in the left cerebellum and less activation in the right lateral orbitofrontal cortex (OFC) and the right dorsolateral prefrontal cortex (DLPFC). When the MJ group was divided into Moderate (8-35 joints/week) and Heavy users (53-84 joints/week), the Heavy MJ group showed less activation in the left medial OFC and greater activation in the left cerebellum than the Moderate group. similar between the Moderate MJ users and Control… very heavy users of MJ have persistent decision-making deficits and alterations in brain activity
Bonnet A, Marchalant Y. 2015. “Potential therapeutic contributions of the endocannabinoid system towards aging and Alzheimer’s Disease” Aging Dis 6(5): 400-405
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4567222/
Alzheimer’s disease (AD) is the most common neurodegenerative disease and accounts for the majority of diagnosed dementia after age 60. It is estimated to currently affect between 20 and 30 million people worldwide with an incidence of the disease between 3 and 30% over the age of 60
Currently available drugs do not reverse or stop the progression of the disease
important feature of AD’s pathology is the presence of a chronic inflammatory component
A lot of the studies have focused on microglial cells, but it is now clear that microglial cells are assisted by astrocytes and endothelial cells [18] to maintain a chronic inflammatory state of the brain
elevated levels of inflammatory factors (such as IL-6 and C-reactive protein notably) could be found in the plasma of AD patients long before clinical onset of the disease [23,24]
Several reports over the last two decades demonstrate the potential benefits from the modulation of the ECS over amyloid β and Tau hyper-phosphorylation
most studies looking at the modulation of neuroinflammation using cannabinoids have mainly focused on CB2 receptors as they are mostly expressed on microglial cells. Studies mentioned earlier [58,68,69,71] have observed reduction in microglial activation and pro-inflammatory cytokines following infusion of Aβ in rodents
ur work demonstrated the anti-inflammatory potential using WIN-55,212-2 chronically on chronic neuroinflammation in young and aged rats [54,56,77] as well the neurogenic and cognitive effect of WIN-55,212-2 in aged animals [55]. Our data suggested that the anti-inflammmatory effect of WIN-55,212-2 could be due to its activity on TRPV1 receptors and its neurogenic effects linked to both CB1 and CB2 receptor
Boos H, Aleman A, Cahn W, Hulshoff H, Kahn R. 2007. “Brain volumes in relatives of patients with schizophrenia” Arch Gen Psychiatry 64:297-304
http://www.ncbi.nlm.nih.gov/pubmed/17339518 – in dropbox
smaller brain volumes in schizophrenia, with more pronounced reductions in the hippocampus and amygdala
estimated heritability of 80% including interaction between the genes and environment
This meta-analysis integrated the results of 25 MRI stud-ies that compared brain volumes of 1065 nonpsychotic first-degree relatives of patients with schizophrenia with those of 1100 healthy control subjects. The results indicate that brain volumes in relatives of patients with schizophrenia differ from those of healthy control subjects, with effect sizes in the small to moderate range. The largest effect is found in hippocampal volume (d = 0.31), with relatives of patients having smaller volumes than healthy control subjects. In addition, total gray matter volume (d = 0.18) and third-ventricle (d = 0.21) volume are smaller in relatives compared with healthy control subjects. Although total brain and white matter volume did not differ significantly in relatives compared with healthy controls, both structures showed a trend toward significance (P = .06 and P = .07, respectively). The analysis that compared patients with schizophrenia with first-degree relatives showed smaller hippocampal volumes in the patients (d = 0.43)
brain abnormalities in schizophrenia are related (in part) to the risk of developing the disease and that these brain changes may therefore predate the clinical onset of the disorder
lower performance in relatives of patients compared with healthy control sub-jects was reported on several cognitive domains, includ-ing verbal and declarative memory, executive function-ing, and attention. Interestingly, Sitskoorn et al 62 found that the largest effect size was obtained for verbal memory (d = 0.54), being significantly worse in relatives of pa-tients than in healthy subjects
decreased verbal memory is one of the most robust neuropsychological findings in schizo-phrenia. 60 Deficits in verbal memory have generally been associated with smaller (left) hippocampal volume, 65 as is also the case in patients with schizophrenia 66,67 and their relatives. 17,26 In the present meta-analysis, the effect size was considerably larger for the left than for the right
hippocampus. This finding is consistent with findings from lesion and functional MRI studies in healthy subjects, suggesting more involvement of the left hippo-campus in encoding and recognition of verbal as opposed to visual or pictorial material
Obstetric complications such as hypoxia are known to result in smaller brain vol-umes, affecting the hippocampus profoundly. 21,72,73 Smaller hippocampal volumes have also been associated with brain injury 65,74,75 and stress 65,76 and have been found not only in schizophrenia but also in several other psychiatric dis-orders, such as major depression, posttraumatic stress dis-order, obsessive-compulsive disorder, and borderline per-sonality disorder. 65 An important function of the hippocampus and amygdala is the regulation of the hy-pothalamic-pituitary-adrenal axis, which plays a role in stress processing. This regulation may be altered be-cause of a genetic predisposition. In depression, the hy-pothalamic-pituitary-adrenal axis is strongly activated and
the adrenal cortex hypersecretes glucocorticoids such as cortisol. Although less pronounced, considerable hypo-thalamic-pituitary-adrenal activation is also found in schizophrenia. 77 On the basis of earlier animal experi-ments, overexposure to cortisol during prolonged peri-ods of stress is expected to damage the brain, particu-larly the hippocampus. Sapolsky et al 77 provided evidence in rats that chronic stress, with the concomitant in-crease in corticoid levels, causes loss of neurons in the hippocampus and subsequent deficits in memory func-tion and cognition. In patients with depression, this glu-cocorticoid cascade has also been presumed to result in decreased hippocampal volume, 78 possibly explained by apoptosis. 79 Both apoptosis and neurogenesis have been shown to occur in the hippocampus. 80 Thus, smaller hip-pocampal volumes in patients with schizophrenia and their first-degree relatives might also be the result of stress-related processes in the brain. 81
slightly elevated rates of obstetric complications are found not only in patients with schizophrenia but also in their nonpsychotic first-degree relatives. 82,83 As reported by Can-non et al,
83 most of these relatives exposed to obstetric complications did not develop schizophrenia
Van Erp et al 9 examined siblings of pa-tients with schizophrenia and found that hippocampal volumes differed stepwise with each increase in genetic predisposition to schizophrenia and that hippocampal vol-umes of patients exposed to fetal hypoxia were smaller than those who were unexposed, whereas no such rela-tionship was observed within the healthy control sub-jects. They suggested that carrying susceptibility genes for schizophrenia makes one vulnerable to perinatal dam-age, especially in the hippocampus
structures other than those that have been evaluated in this meta-analysis may also be affected in relatives of patients with schizophrenia
the possibility that some of the effects found in this meta-analysis were caused by confounding factors such as sex and age cannot be ruled out
Borgwardt S, McGuire P, Aston J, Gschwandtner U, Pfluger M, Stieglitz R-D, Radue E-W, Riecher-Rossler A. 2008. “Reductions in frontal, temporal and parietal volume associated with the onset of psychosis” Schizophrenia Research 106: 108-114
http://www.ncbi.nlm.nih.gov/pubmed/18789654 add to dropbox
Which volumetric abnormalities are related to psychotic illness, as opposed to vulne rability to psychosis is unclear
n = 20). Over the following 3 years, 10 subjects developed psychosis and 10 did not. Subjects were re-scanned af ter the ons et of psychosis or at the end of follow-up if they did not become psychotic… In subjects who developed psychosis there were longitudinal volume reductions in the orbitofrontal, superior frontal, inferior temporal, medial and superior parietal cortex, and in the cerebellum
longitudinal reductions in regional gray matter volume also occur in chronic patients ( Cahn et al., 20 02; Ho et al., 20 03; Kasai et al., 20 03; Kubicki et al., 20 02; Mathalon et al., 20 01; Sporn et al., 20 03; Wood et al., 20 01)
Bossong M, Berckel B, Boellaard R, Zuurman L, Schuit R, Windhorst A, van Gerven J, Ramsey N, Lammertsma A, Kahn R. 2009. “Δ9-Tetrahydrocannabinol Induces Dopamine Release in the Human Striatum” Neuropsychopharmacology 34: 759-766
http://www.nature.com/npp/journal/v34/n3/full/npp2008138a.html
Miniscule finding among general absence of finding exaggerated and made to sound like proof of addictiveness, because Bossong is a lying weasel of a scientist
“This is the first study demonstrating THC-induced dopamine release in the human striatum… The ability of THC to induce dopamine release in the human striatum suggests that THC shares addictive properties with other drugs of abuse.”
whereas amphetamine (Drevets et al, 2001;Martinez et al, 2003, 2007), cocaine (Schlaepfer et al, 1997), alcohol (Boileau et al, 2003), and nicotine (Brody et al, 2004, 2006) cause reductions in dopamine D2/D3receptor availability in the range of 10–30%, we found a relatively modest THC-induced decrease of 3.4 and 3.9% in the ventral striatum and the precommissural dorsal putamen
THC inhalation reduces [11C]raclopride binding in the ventral striatum and the precommissural dorsal putamen but not in other striatal subregions. This is consistent with an increase in dopamine levels in these regions
As THC was dissolved in 100 vol% alcohol and the solvent was used as placebo, we cannot exclude that the inhalation of alcohol has caused dopamine release. However, this is very unlikely, as only 200 μl alcohol was administered
THC plasma concentrations in this study were comparable with or even higher than those obtained after smoking of high-potency cannabis
it is unlikely that striatal dopamine release immediately after THC administration has been much larger than the modest levels of 3.4–3.9%
Thus, elevated striatal dopamine release after the use of cannabis may explain how cannabis use contributes to the development and pathophysiology of schizophrenia
in animals, it has been demonstrated that cannabinoid substances such as Δ9-tetrahydrocannabinol (THC), the main psychoactive component in cannabis (Gaoni and Mechoulam, 1964), also stimulate striatal dopamine neurotransmission (for review see Tanda and Goldberg, 2003; Lupica et al, 2004; Gardner, 2005). Cannabinoids enhance neuronal firing of mesolimbic dopamine neurons (French, 1997; French et al, 1997; Gessa et al, 1998) and elevate striatal dopamine levels (Ng Cheong Ton et al, 1988; Chen et al, 1990; Tanda et al, 1997; Malone and Taylor, 1999; Fadda et al, 2006), both through activation of cannabinoid CB1 receptors (French, 1997; French et al, 1997;Tanda et al, 1997; Gessa et al, 1998; Malone and Taylor, 1999). However, whether THC affects the human striatal dopamine system is currently unknown
Nine healthy male subjects gave informed consent for this study. Seven volunteers completed the study procedure. One subject was excluded because of positive urine drug screening on the first study day. Another subject did not complete the second scanning session because of anxiety… All subjects were familiar with the effects of cannabis. Two subjects used cannabis less than once a month, one subject three times a month, three subjects twice a month and one subject used cannabis once a week. They all showed negative urine screening at both study days
There were no significant changes in equilibrium levels of striatal activity, expressed by percentage change of activity concentration over time, between placebo (−0.10±0.16% per min) and THC (−0.12±0.08% per min) sessions (p=0.633). In addition, the slope (ie change over time during scanning) of the ratio between striatum and cerebellum [11C]raclopride concentrations was not significantly different between sessions (−0.0011±0.0030 and −0.0010±0.0025 for placebo and THC sessions, respectively; p=0.967)
[11C]Raclopride concentrations were not significantly different between placebo and THC sessions in both whole blood (0.59±0.25 and 0.61±0.28 kBq/g, respectively;p=0.717) and plasma (1.02±0.42 and 1.06±0.43 kBq/g, respectively; p=0.687), normalized to the effectively injected dose. In addition, the fraction of parent[11C]raclopride was not significantly different (p=0.191) between placebo (80.4±5.6%) and THC (77.5±4.3%).
significantly reduced in the ventral striatum and the precommissural dorsal putamen after inhalation of THC compared to placebo
No significant associations between measures of dopamine release and behavioral, subjective or physiological effects were demonstrated.
Bossong M, Niesink R. 2010. “Adolescent brain maturation, the endogenous cannabinoid system and the neurobiology of cannabis-induced schizophrenia” Progress in Neurobiology 92: 370-385
http://www.ncbi.nlm.nih.gov/pubmed/20624444 in dropbox
Central argument of the paper is based on the assumption that marijuana increases glutamate signalling, for which no evidence is provided, and evidence that it does the opposite is brushed off, because Bossong is a lying weasel of a scientist
implies a toxic substance affecting the central nervous system during a critical period, resulting in irreversible structural changes. These changes subsequently cause psychopathological effects. It is assumed that D 9-tetrahydrocannabinol (D 9-THC, henceforth mentioned as THC), the main psychoactive substance in cannabis, is the neurotoxic substance and that adolescence is the critical period. The interference of THC with a maturational process in the brain of adolescents is supposed to induce structural and functional changes.
From the results of the literature search, it can be postulated that THC adversely affects normal physiological maturational processes during adolescence. Usually, the interaction of endoge-nous cannabinoids with the CB1 receptor is critically involved in brain maturation through its regulating role in the release of glutamate. Through its action on CB1 receptors, THC can interfere
with this normal physiological process, resulting in disturbed glutamate release, subtle neurotoxic effects and subsequent structural defects. Since maturation of the prefrontal cortex (PFC) is one of the most important processes during adolescence, THC may predominantly affect the maturation of specific neurocircuitries within this brain region
Brain development is an organized and highly dynamic multistep process, which is genetically determined, epigenetically directed and environmentally influenced (Tau and Peterson, 2010)
From childhood to adolescence, development shifts from producing a large number of neurons to creating efficient neuronal pathways. This efficiency is thought to be achieved by synaptic refinement, the process by which some connections between brain cells are pruned and eliminated, and ‘‘useful’’ neurons, synapses and dendrites are selected and preserved for the adult brain (Katz and Shatz, 1996; Cohen-Cory, 2002; Whitford et al., 2007; Purves et al., 2008; Luna, 2009)
Recent studies focusing on the development of the social brain support evidence that adolescence also represents a period of significant social development
For information from the outside world, cortical areas sub-serving high-order behavior completely depend on subcortico-cortical connections. Animal studies, specifically studies in rodents, suggest unique anatomical and functional changes during adolescence. An intact innervation of the cortex from subcortical structures during this time period appears to be a prerequisite for the proper maturation of specific cortical areas. Thus, lesions in neurons connecting subcortical structures, such as the amygdale and hippocampus, with the prefrontal cortex before adolescence lead to specific structural and behavioral changes at the end of adolescence and in adulthood. Such lesions studies have been proposed as good animal models for neurodevelopmental psycho-pathological disorders, such as schizophrenia (Bouwmeester et al., 2002; Lipska, 2004; Tseng et al., 2009 )
Deprivation of complex stimuli in vulnerable periods, such as rearing in an impoverished environment ( Benefiel et al., 2005 ) and social isolation ( Hall, 1998), results in permanent disturbances in adult behavior, disturbances that seem to be mediated by defects in the neural circuitry within the PFC (Card et al., 2005 ). Deprivation studies also have shown that social play behavior, the first form of non-mother directed social behaviors displayed by most adolescent mammals, is essential for appropriate social, cognitive and sexual development ( Vanderschuren et al., 1997)
the development of complex functions involves cascades of sensitive periods affecting different levels of processing at different ages (Knudsen, 2004). Although stimuli from the cognitive, social and emotional domain are thought to be important for appropriate brain maturation during adolescence, it does not exclude an important role for this type of stimuli during critical or sensitive periods during infancy and childhood. For example, social contacts with parents has shown to be important for brain plasticity early in life (Helmeke et al., 2009; Musholt et al., 2009), whereas social contacts with peers are thought to be important for brain development during adolescence (Leussis and Andersen, 2008)
Transient blockade of the CB1 receptor, for example by exposure to exogenous cannabinoids such as THC (1), disrupts the protective effect of the endogenous cannabioid system (2), thereby causing an excess of glutamate (3) and consequently too great an influx of Ca2+ (4) in the postsynaptic neuron. This causes a disturbance of the LTP/LTD balance, which may lead to pruning of the postsynaptic part of the synapse and possibly of the postsynaptic dendritic arbors. The ultimate result might be a disturbance in local neuronal circuitry
overactivation of ionotropic glutamate receptors can induce either apoptosis or necrosis through excessive Ca2+ influx, a process known as excitotoxicity (Mody and MacDonald, 1995; Olney, 2003). This is illustrated by the fact that the immature brain is more sensitive to the excitotoxic effects of glutamate than the adult brain ( Ikonomidou et al., 1999; Olney et al., 2000 ). Because the immature NMDA receptor is more sensitive to glutamate, the probability to induce such excessive concentrations of postsynap-tic intracellular Ca2+ is increased during the critical period. Excitotoxicity may disturb the normal maturational processes by the formation of aberrant cortical connections (Olney, 2003). Thus, binding of glutamate to the NMDA receptor will have greater consequences during the critical period than outside this phase.
A comprehensive neuroanatomical analysis of CB1 receptors and endogenous cannabinoids during specific critical periods within particular areas of the frontal cortex is currently lacking
The research that has been done in vulnerable people concerns studies in people diagnosed with subclinical symptoms ( Henquet et al., 2004 ). In fact, these people already suffered from a subclinical prodromal syndrome at the time that the exposure to cannabis took place
in this model, a dose–time–effect relationship is responsible for the toxic effect of cannabis during late postnatal development. This is possibly also one of the main reasons why not everyone who used cannabis during adolescence will develop a permanent psychosis. Lower exposure will result in a less serious, probably sub-threshold effect
Most of the research on the neurobiological basis of schizophrenia is monodisciplinary and the same applies to the study of the action and function of endogenous cannabinoids, although both require a multidisciplinary approach
Bossong M, van Hell H, Jager G, Kahn R, Ramsey N, Jansma J. 2013. “The endocannabinoid system and emotional processing: a pharmacological fMRI study with Δ9-Tetrahydrocannabinol” European Neuropsychopharmacology 23(12): 1687-1697
http://www.europeanneuropsychopharmacology.com/article/S0924-977X(13)00195-8/fulltext in dropbox
THC) on brain function related to emotional processing in 11 healthy subjects… Performance and brain activity during matching of stimuli with a negative (‘fearful faces’) or a positive content (‘happy faces’) were assessed after placebo and THC administration. After THC administration, performance accuracy was decreased for stimuli with a negative but not for stimuli with a positive emotional content
These results suggest that administration of THC shifts the brain’s bias for stimuli that have a negative impact towards a bias for stimuli that have a positive impact.
a defect in endocannabinoid neurotransmission could contribute to the abnormal emotional reactions as seen in patients with a major depression
As emotional responses such as anxiety and fear are associated with increased glutamate and diminished GABA neurotransmission (Millan, 2003), the reduced negative emotional bias as demonstrated in the current study may be the result of a THC-induced reinstatement of the balance between both neurotransmitter systems (Ruehle et al., 2012)
We did not find significant effects of THC on amygdala activity. Possibly, the subjective anxiety-like effects of THC administration may have specifically masked THC-induced effects on the response of the amygdala, as it has been shown that particularly amygdala activity may be involved in the subjective response to pharmacologically induced anxiety (Eser et al., 2009). This view is supported by results of Fusar-Poli et al. (2009)), who showed strong subjective anxiety-like effects of THC, but no significant effects of THC administration on the amygdala response
Bossong M, Jansma J, van Hell H, Jager G, Kahn R, Ramsey N. 2013. “Default mode network in the effects of Δ9-Tetrahydrocannabinol (THC) on Human Executive Function” PLoS One 8(7): e70074
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3729458/
Decreases in contentedness and calmness may be related to THC administration in an MRI environment, as they were not shown in a previous study using Positron Emission Tomography (PET) [34]
THC-induced impairment of task performance as shown in the present study is expected to result in increased effort to maintain good performance levels, most likely reflected as elevated CES activity. However, THC did not affect CES activity during performance of the task.
Previous studies have reported reduced activity in the CES in psychiatric disorders such as ADHD [58],[59] and schizophrenia [60], [61], an effect that is likely related to impaired task performance [61]. One explanation for the apparent discrepancy with the current findings could be that performance deficits as shown in our study after THC are moderate compared to those of psychiatric patients. For example, decreased CES activity in schizophrenia patients in the study of Salgado-Pineda et al. [60] was associated with a 33% reduction in the mean percentage of correctly identified targets
Higher COMT activity, resulting in reduced prefrontal dopamine neurotransmission, has been associated with significantly greater deactivation of the posterior cingulate cortex of healthy volunteers [64] and reduced deactivation in the medial prefrontal cortex of both healthy subjects and schizophrenia patients … Interestingly, individuals with increased COMT activity appear to have stronger responses to THC administration in terms of acute psychotic effects and cognitive impairments [67]
nicotine administration decreased DMN activity at rest in non-smokers [70], improved cognitive withdrawal symptoms of abstinent smokers through modulation of functional connectivity within the DMN and of inverse coupling between default mode and central executive brain networks [71], and enhanced visuospatial attention by deactivating DMN nodes including posterior cingulate cortex and angular gyrus in minimally deprived smokers [15]
neither brain activity nor behavioral effects were significantly correlated with reported cannabis use (data not shown)
non-specific THC-induced changes on cerebral blood flow may have confounded our results [72]
this study shows that THC administration results in less deactivation in the DMN during an executive function task, an effect that is correlated with task performance. These results suggest an important role for the eCB system in both DMN modulation and executive function. The association of the eCB system with DMN modulation may be relevant for psychiatric disorders associated with executive function deficits, such as schizophrenia and ADHD, as well as for neurological disorders such as Alzheimer’s disease
Bossong M, Jansma J, Bhattacharyya S, Ramsey N. 2014. “Role of the endocannabinoid system in brain functions relevant for schizophrenia: an overview of human challenge studies with cannabis or delta9-tetrahydrocannabinol (THC)” Progress in Neuro-Psychopharmacology & Biological Psychiatry 52: 53-69
http://www.ncbi.nlm.nih.gov/pubmed/24380726 in dropbox
Pattern in text of “Schizophrenia involves significant impairment of X; studies showed THC does not impair X, but alters activity during the task in a strikingly different pattern than the alterations in schizophrenia; altogether they show similarity” My working hypothesis remains that Bossong is a lying, weasel of a scientist. X = increase in dopamine signalling, memory impairment, executive function, emotional processing, response inhibition, auditory and visual processing. How does this bullshit get published?
“administration of… THC… reduces most cognitive functions” (Bossong is a lying weasel of a scientist)
the incidence of schizophrenia is relatively low (10-22 per 100,000), its prevalence is relatively high (0.3-0.7 per 100) due to the chronic nature of the illness (McGrath, 2008)
One of the most robust pathophysiological features of schizophrenia is an increase in striatal dopamine function. … dopamine enhancing drugs are able to induce direct psychotic effects (Angrist and van Kammen, 1984; Lieberman et al, 1987)… PET and SPECT neuroimaging sturides have consistently shown increased dopamine neurotransmission and elevated dopamine synthesis capacity in the striatum of patients with schizophrenia (for reviews Abi-Dargham, 2004; Howes 2009)
THC induces strong behavioral and physiological effects which are not associated with a large magnitude of dopamine release in the striatum
cognitive impairments are considered a core feature of the illness (Elvvag and Goldberg 2000; Gold 2004; Green 1996)
in two extensive meta-analyses, most prominent deficits during encoding of information were found in prefrontal areas of patients, including right medial, superior and middle frontal gyrus, as well as bilateral inferior frontal gyrus (Achim and Lepage 2005; Ragland 2009)
areas that showed significantly increased activity in schizophrenia patients during encoding included left precentral gyrus, middle temporal gyrus and postcentral gyrus, right parahippocampal gyrus, and anterior cingulate (Ragland, 2009), suggesting that general deficits in cognitive control may strongly contribute to memory impairment in schizophrenia (Andrews 2006; Ragland 2009; Segarra 2008)
Many neuropsychological studies investigating the impact of a challenge with cannabis or THC on memory paradigms consisting of encoding and recall conditions have shown an absence of effect, despite robust subjective and physiological responses (Block, 1984; Chait, 1994; Darley, 1977; Hart, 2001,2002, 2010; McDonald, 2003). Recall of information learned before cannabis use is also generally not affected (Abel, 1971; Darley, 1973; Dornbush, 1974). However, effects of cannabis or THC on memory performance have been reported in the free recall of information that is previously learned under the influence of cannabinoids (Curran, 2002; D’Souza, 2004; Liem-Moolenaar, 2010; Miller, 1978; Miller, 1976; Miller, 1977a,b; Miller 1979; see for reviews Ranganathan and D’Souza; Schoeler and Bhattacharyya, 2013; Zuurman, 2009). This suggets that cannabinoids may specifically influence encoding but not recall of information
a normal decrease in activity in the parahippocampal gyrus that was present over repeated encoding blocks during placebo was no longer evident after THC administration. THC also eliminated the linear reduction in activity in the anterior cingulate and bilateral medial prefrontal cortex that was shown over recall conditions. … overall task performance was unaffected… THC also attenuated bilateral striatal activity across repeated recall blocks, and its effect in the ventral striatum was directly correlated with the severity of THC-induced psychotic symptoms (Bhattacharyya, 2009)
One possibility is that subjects with higher dopamine levels show a more inefficient encoding of information after THC administration, which may be compensated by increased midbrain activity during recall to maintain normal task performance
schizophrenia patients exhibited increases in activity in left pre- and postcentral gyrus and temporal cortex during encoding, and reductions in bilateral thalamus and cerebellum during recall (Achim and Lepage, 2005; Ragland, 2009), effects that were not seen after THC administration [followed by some irrelevent excuses of why differences may exist]
Deficits in working memory, which is the ability for short-term storage and manipulation of information (Baddeley, 1986) are a persistent and disabling feature of schizophrenia that may underlie many cognitive impairments and symptoms… schizophrenia patients often exhibit a reduced load-dependent increase in brain activity, together with enhanced brain activity for tasks with low working memory load. This has been shown in a network of working-memory related brain areas, including bilateral dorsolateral prefrontal cortex, inferior parietal gyrus, basal ganglia, thalamus and anterior cingulate (Callicott 2000; Jansma 2004; Johnson 2006; Manoach 1999, 2000; Potkin 2009)
THC administration enhanced activity for low working memory loads, and reduced the linear relationship between working memory load and activity in a network of working-memory related brain regions and in the left dorsolateral prefrontal cortex, inferior temporal gyrus, inferior parietal gyrus and cerebellum… performance accuracy after THC was only reduced for oderately high working memory loads
working memory inefficiency was demonstrated in the left cerebellum and inferior temporal gyrus after THC administration but not in schizophrenia patients… differences in study design most likely account for the variation in results
Neuroimaging studies with schizophrenia patients have shown reduced activity in the central executive system, mainly in prefrontal and parietal areas, during perfromance of executive function tasks such as the CPT (Gur, 2007; Hong, 2011; Kiehl, 2005; Salgado-Pineda, 2004; Volz, 1999; Weiss, 2007), and effect that was related to impaired task performance
schizophrenia patients exhibit an inability to deactivate the default mode network during performance of executive function paradigms (Garrity, 2007; Hasenkamp, 2011; Pomarol-Clotet, 2008; Schneider, 2011; Whitfield-Gabrieli, 2009; see for reviews Broyd, 2009; Whitfield-Gabrieli and Ford, 2012)
A number of neuropsychological studies have reported no effects of cannabis or THC administration on continuous performance paradigms (Casswell, 1975; Curran, 2002; D’Souza, 2004; Vachon, 1974; Weil, 1968; Wilson, 1994)… decreased performance after cannabis or THC has been reported on more challenging continuous performance paradigms (Hunault, 2009; Sharma and Moskowitz, 1974). Tasks used in these studies are difficult cognitive tasks with a low memory component, typically relying on fast processing of information
Bossong 2013: task performance impaired after THC administration, reflected in both an increase in false alarms and a reduction in detected targets, associated with reduced deactivation in a set of brain regions linked to the default mode network, including posterior cingulate cortex and angular gyrus, correlated with lower performance after THC. Regions that were activated by the continuous performance task, notably bilateral prefrontal and parietal cortices, did not show effects of THC. [in the study, it was noted that the effect was moderate compared to psychiatric subjects, subjects given THC were less calm and more distracted by the fMRI environment, and that changes in cerebral blood flow likely confounded the results, none of which is mentioned here]
Reduced performance on executive function paradigms [NOT ESTABLISHED] after cannabis or THC may be associated with a failure to deactivate teh default mode network rather than effects on activity in the central executive system (Bossong, 2013). … resting state fMRI study showed that THC administration decreased functional connectivity between the posterior cingulate cortex and a network of brain regions collectively referred to as the left dorsal visual stream, thought to be involved in attentional processes (Klumpers, 2012). These findings are in line with impairments in capacity to deactivate the default mode network during performance of executive function paradigms as exhibited by schizophrenia patients (Broyd, 2009; Whitfield-Gabrieli and Ford, 2012)
Acute THC administration did not affect brain activity patterns in the central executive system… in contrast with reduced activity in the central executive system of schizophrenia patients during performance of executive function tasks (Gur, 2007; Salgado-Pineda, 2004).
One explanation for this apparent discrepancy could be that performance deficits as shown in the study by Bossong and colleagues (9% decrease in the mean percentage of correctly identified targets) are moderate compared to those of schizophrenia subjects. For example, decreased activity in the central executive system of schizophrenia patients in the study of Salgado-Pineda (2004) was associated with a 33% reduction in the mean percentage of correctly identified targets. Another possibility is that the ability to detect erroneous responses is impaired in schizophrenia patients, but not after THC administration, as diminished capacity for error monitoring is associated with reduced activity in the central executive system (Klein, 2007; Ridderinkhof, 2004)
Abnormalities in emotional processing are among the most important characteristics of schizophrenia, with significant consequences for social functioning and subjective well-being of patients (Aleman and Kahn, 2005; Pinkham, 2003). Individuals with schizophrenia have deficits in facial affect perception, which are present throughout the course of the disorder, and which are stronger for the perception of negative than positive emotions (Edwards, 2002; Mandal, 1998; Pinkham, 2003)… a large network of brain regions regulates processing of emotions, including [amygdala], orbital frontal cortex, prefrontal cortex, anterior cingulate, and occipital and temporal lobes (Fusar-Poli, 2009; Phillips, 2008). Schizophrenia patients exhibit reduced activity in this network when emotional stimuli are compared to similar but neutral stimuli (Gur, 2002; Aleman and Kahn, 2005) which is due to increased activity to neutral stimuli in schizophrenia (Hall, 2008; Holt, 2006; Surguladze, 2006). … schizophrenia patients activate areas related to processing of emotions in response ton netural stimuli to a stronger extent than normal subjects, consistent with the theory that schizophrenia may be characterized as a state of aberrant assignment of salience to otherwise insignificant stimuli (Kapur, 2003).
Only one neuropsychological study examined the acute effects of THC on processing of emotions (Ballard, 2012)… THC significantly reduced recognition of threatening stimuli (fear and anger), but did not affect ratings of emotional scenes… reduced activity in amygdala to threatening stimuli with no difference on task performance found by Phan, 2008.
When threatening stimuli were compared to neutral stimuli, THC administration increased activity in the left precuneus and reduced activity in the right inferior frontal gyrus, right superior temporal gyrus and left medial frontal gyrus, without effects on task performance (Fusar-Poli, 2009b)… in 2010 failed to show any effects of THC administration on effective connectivity etween the amygdala and the anterior cingulate cortex
studies in unmedicated schizophrenia patients have consistently demonstrated attenuated activity in the ventral striatum during reward anticipation (Esslinger, 2012; Juckel, 2006; Nielsen, 2012a; Schlagenhauf, 2009)
Eleven recreational cannabis users showed similar behavioral resposnes to reward during THC and placebo… THC attenuated the brain response during reward feedback in a network of areas involved in the processing of feedback including the bilateral inferior temporal gyrus, left inferior parietal gyrus and right superior frontal gyrus. Brain activity during the anticipation of reward was not affected. (van Hell, 2012)
Jansma (2013) compared activity in the ventral striatum and the caudate putamen during anticipation and feedback of reward after THC inhalation (6 mg) between subjects with a nicotine addiction (N=10) and healthy controls (N=11) (matched on cannabis use). In the absence of any other differences between the two groups, THC administration induced a significantly different effect on activity in the ventral striatum during reward anticipation: subjects with a nicotine addiction exhibited a significant reduction in activity after THC, not present in healthy controls
Schizophrenia has been associated with impaired response inhibition for decades (Frith, 1979)… attenuated activity in patients known to be involved in this cognitive task including bilateral dorsolateral prefrontal cortex (Arce, 2006; Ford, 2004; Rubia, 2001), anterior cingulate cortex (Arce, 2006; Rubia, 2001), right inferior frontal gyrus (Kaladjian, 2007), bilateral superior and inferior parietal gyrus (Ford, 2004) and right striatum (Vink, 2006)… abnormalities also in bilateral superior temporal gyrus (Ford, 2004)
In the absence of THC-induced effects on reaction time or inhibition errors, activity was reduced in right inferior frontal gyrus, bilateral anterior cingulate gyrus and precuneus, and increased in right (para)hippocampal gyrus, right superior temporal gyrus and left posterior cingulate gyrus (Borgwardt, 2008). In an extension of this study, THC showed significantly stronger [than absent?] effects on inhibition errors in the group of participants with psychotic symptoms, accompanied by different effects of THC between groups on activity in the left parahippocampal gyrus, bilateral middle temporal gyrus and right fusiform gyrus (increased in right middle temporal, decreased in other areas) (Atakan, 2013)
The aberrant processing of salience is thought to be a fundamental factor underlying psychosis (Kapur, 2003)… patients exhibited a diffuse pattern of decreased activity during processing of novel auditory stimuli, with the strongest reductions in bilateral inferior parietal lobe, bilateral caudate/thalamus, left postcentral gyrus, right posterior cingulate gyrus and left inferior temporal gyrus (Kiehl, 2005). Gur (2007) showed decreased activity in bilateral cuneus and left middle occipital gyrus, but increased activity in bilateral inferior parietal lobule, right middle and inferior frontal gyrus, and left fusiform gyrus during visual processing of novel stimuli. Jensen (2008) showed increased activity in right ventral striatum, middle cingulate cortex, thalamus, prefrontal cortex and hippocampus in response to neutral condition in motivational salience task
THC administration reduced the response latency to standard stimuli relative to oddball stimuli… attenuated activity in right caudate and putamen, increased activity in right prefrontal cortex during oddball processing. Effect of THC in right caudate was negatively correlated with both the severity of THC-induced psychotic symptoms and the effect on response latency
Hallucinations are one of the main clinical features of schizophrenia and can occur in any sensory modality, although auditory and visual hallucinations are most common (Allen, 2008). Elevated activity in the auditory and visual processing pathways has consistently been shown when patients are experiencing auditory and visual hallucinations, respectively (Allen, 2008; Bossong and Allen, in press). In response to external speech, patients exhibit strong reductions in activity in the superior temporal gyrus, althouh decreases in other area (bilateral middle frontal gyrus, right insula, left putamen) and increases (right middle temporal gyrus, left inferior parietal lobule, anterior cingulate cortex) have also been reported (Copolov, 2003; Woodruff, 1997)
THC decreased activation relative to placebo in bilateral superior temporal gyrus and right middle temporal gyrus when subjects listened passively to words read. The attenuation of temporal activation by THC was associated with increase in psychotic symptoms. During viewing of a visual checkerboard, THC reduced activity in regions of the occipital lobe (left middle occipital gyrus, right cuneus, bilateral lingual gyrus), increased in right middle occipital gyrus, bilateral lingual gyrus (Winton-Brown, 2011)
“Similarities in brain function between healthy volunteers after THC administration and schizophrenia patients provide an argument for the role of the endocannabinoid system in symptoms of schizophrenia”
Significant decreases in alertness may be associated with activity changes related to cognitive brain functions discussed in this review paper
Bossong M, Mehta M, van Berckel B, Howes O, Kahn R, Stokes P. 2015. “Further human evidence for striatal dopamine release induced by administration of ∆9-tetrahydrocannabinol (THC): selectivity to limbic striatum” Psychopharmacology (Berl) 232(15):2723-9
http://www.ncbi.nlm.nih.gov/pubmed/25801289
THC administration induced a significant reduction in [(11)C]raclopride binding in the limbic striatum (-3.65 %, from 2.39 ± 0.26 to 2.30 ± 0.23, p = 0.023). This is consistent with increased dopamine levels in this region. No significant differences between THC and placebo were found in other striatal subdivisions
This finding suggests limited involvement of the endocannabinoid system in regulating human striatal dopamine release and thereby challenges the hypothesis that an increase in striatal dopamine levels after cannabis use is the primary biological mechanism underlying the associated higher risk of schizophrenia.
[MV: I’d previously accused Bossong of dishonesty for claiming that marijuana has effects on dopamine “similar to other drugs of abuse,” even though his results show the effect to be smaller by an order of magnitude. Finally he admits that this effect cannot explain the association with psychosis. If he will also admit that THC does not block cannabinoid receptors or produce elevated glutamate signalling, I might take him off my shit list. His speculations have emboldened anti-pot critics despite the lack of empirical support. Here he finally admits a lack of empirical support for at least one part of that]
Boulanger L. 2009. “Immune proteins in brain development and synaptic plasticity” Neuron 64(1): 93-109
http://www.sciencedirect.com/science/article/pii/S0896627309006783
The MHCI is a large family of proteins that are expressed on the surface of most nucleated cells in the body. Cell-surface MHCI presents peptides derived from intracellular proteins for immune surveillance, permitting immune recognition of foreign (“nonself”) antigens generated by transplanted, infected, and cancerous tissues
Neurons themselves express immune proteins, but they can also be affected by secreted and cell-surface immune proteins produced by infiltrating immune cells (e.g., lymphocytes), microglia (the resident CNS macrophage), and other resident neuroglia (e.g., astrocytes, oligodendrocytes). Indeed, although neurons can produce tumor necrosis factor- (TNF)- α, glia are the source of the endogenous TNFα that affects synaptic scaling
Interleukin- (IL-) 6 and IL-6 receptor mRNAs are also coexpressed in neurons and are developmentally regulated in rat brain, with highest levels of both detected in adult hippocampus (Gadient and Otten, 1994)
TNFα can promote the cell surface accumulation of AMPA-type glutamate receptors (AMPARs) in hippocampal neurons in vitro. Since AMPARs carry the majority of excitatory glutamatergic current at resting membrane potentials, this TNFα-mediated increase in cell surface AMPARs should regulate synaptic transmission, and indeed, exogenous TNFα rapidly induces an increase in the frequency and amplitude of miniature excitatory postsynaptic currents (mEPSCs) in hippocampal neurons in vitro (Beattie et al., 2002 and Stellwagen et al., 2005)
In addition to its regulation of the properties of excitatory connections, MHCI was recently identified in a screen for targets of the transcription factor Npas4, which controls the number of inhibitory GABA-releasing synapses that contact excitatory neurons (Lin et al., 2008). However, it remains unknown if MHCI affects the establishment, maintenance, or function of GABAergic connections. A number of proinflammatory cytokines, including IL-1β (Zeise et al., 1992) and TNFα (Stellwagen et al., 2005), have been shown to regulate inhibitory synaptic transmission
endogenous MHCI in the adult hippocampus inhibits LTP and either permits or promotes LTD. A component of many MHCI immunoreceptors, CD3ζ, is also required for LTD and limits LTP in the adult hippocampus (Huh et al., 2000) (Figure 3B)
Since TNFα is a proinflammatory cytokine and can regulate MHCI expression, any effects TNFα may have on plasticity could potentially be mediated by changes in MHCI. In fact, several other proinflammatory cytokines, including interleukin- (IL-) 6 (Bellinger et al., 1995, Li et al., 1997 and Tancredi et al., 2000), IL-1β (Cunningham et al., 1996 and Katsuki et al., 1990), IL-2 (Tancredi et al., 1990), IL-18 (Curran and O’Connor, 2001), IL-8 (Xiong et al., 2003), and interferon-α and -β (D’Arcangelo et al., 1991 and Mendoza-Fernandez et al., 2000), also inhibit hippocampal LTP, consistent with the possibility they may converge on a common pathway. Although exogenous IL-6 inhibits LTP in hippocampal slice, IL-6 levels are dramatically upregulated by LTP induction in vivo (Balschun et al., 2004 and Jankowsky et al., 2000), and application of an anti-IL-6 antibody 90 min after tetanus prolonged LTP and improved long-term memory (Balschun et al., 2004)
In hippocampal neurons in vitro, chronic, long-term reduction of excitatory synaptic activity (e.g., with tetrodotoxin [TTX]) normally causes an increase in synaptic transmission, while a chronic increase in activity (e.g., by application of picrotoxin, a blocker of GABAA-mediated inhibition) causes a decrease in synaptic transmission. These changes keep the activity of networks relatively constant in the face of ongoing acute plasticity, keeping excitation and inhibition in balance and preventing runaway excitation that can lead to epileptic activation and excitotoxicity (Turrigiano, 2008)
MHCI is bidirectionally regulated by changes in activity, with increases in activity adding to the normal constitutive expression of MHCI. In contrast, TNFα may be released in response to a drop in activity ( Stellwagen and Malenka, 2006)
while TNFα is required for scaling up of excitatory synapses, other factors are responsible for scaling them down ( Stellwagen and Malenka, 2006)
Accumulating evidence indicates that the same molecules can perform both immunological and neurological functions in a single organism. It will be important to determine how these distinct functions coexist, and the pressures they exert on one another. Shared molecular machinery could help coordinate neuronal and immune responses of the disparate systems in which these proteins function, linking changes in the timing and magnitude of the two responses. It also increases the risk of potentially pathological molecular crosstalk. Given that the precisely tuned functioning of the immune system and the nervous system are both critical for survival, adaptations may have arisen to minimize such crosstalk, which if unchecked could promote autoimmunity, hamper efforts to fight off infections or cancer, or induce major motor or cognitive impairments. One speculative idea is that the immune privilege of the brain, long taken to be evidence that neurons lack key immune proteins, is instead necessary to preserve the distinct neuronal functions of these proteins. In this model, rather than being evidence of a lack of immune proteins, CNS immune privilege is the opposite: a specialization that is due to the expression and functional importance of immune proteins in normal brain development and plasticity
Bovasso G. 2001. “Cannabis abuse as a risk factor for depressive symptoms” American Journal of Psychiatry 158(12): 2033-2037
http://ajp.psychiatryonline.org/doi/full/10.1176/appi.ajp.158.12.2033
Participants (N=1,920) in the 1980 Baltimore Epidemiologic Catchment Area (ECA) study who were reassessed between 1994 and 1996 as part of a follow-up study
those who reported no depressive symptoms at baseline (N=849) and those with no diagnosis of cannabis abuse at baseline (N=1,837)
In participants with no baseline depressive symptoms, those with a diagnosis of cannabis abuse at baseline were four times more likely than those with no cannabis abuse diagnosis to have depressive symptoms at the follow-up assessment, after adjusting for age, gender, antisocial symptoms, and other baseline covariates. In particular, these participants were more likely to have experienced suicidal ideation and anhedonia during the follow-up period. Among the participants who had no diagnosis of cannabis abuse at baseline, depressive symptoms at baseline failed to significantly predict cannabis abuse at the follow-up assessment
underscore the importance of cannabis abuse prevention rather than treatment
Boys A, Marsden J, Strang J. 2001. “Understanding reasons for drug use amongst young people: a functional perspective” Health Education Research 16(4): 457-469
http://her.oxfordjournals.org/content/16/4/457.full
364 young poly-drug users recruited using snowball-sampling methods (345 cannabis users)
functions most endorsed for cannabis use were to `RELAX’ (96.8%), become `INTOXICATED’ (90.7%), `ENHANCE ACTIVITY’ (72.8%), `DECREASE BOREDOM’ (70.1%), `SLEEP’ (69.6%), FEEL BETTER’ (69.0%) after effects of other substances 64.6, stop worrying 57.7
| Used [substance] to… |
Cannabis (n =345) |
Amphetamines (n = 160) |
Ecstasy (n =357) |
LSD (n =58) |
Cocaine (n =168) |
Alcohol (n =312) |
| Make yourself feel better when down or depressed (FEEL BETTER)a |
69.0 |
43.1 |
48.4 |
20.7 |
61.9 |
69.9 |
| Help you `keep going’ on a night out with friends (KEEP GOING) |
35.9 |
95.6i |
91.1i |
58.6iv |
84.5i |
66.7iv |
| Help you feel elated or euphoric (ELATED/EUPHORIC) |
46.1 |
60.6iv |
77.7iii |
72.4ii |
57.1iv |
51.3 |
| Just get really stoned or intoxicated (INTOXICATED) |
90.7ii |
55.0v |
68.2v |
77.6i |
66.1iii |
89.1i |
| Help you lose weight (LOSE WEIGHT) |
– |
23.1 |
7.0 |
– |
6.0 |
– |
| Help you enjoy the company of your friends (ENJOY COMPANY) |
66.4 |
58.1 |
63.1 |
58.6v |
61.3v |
74.0iii |
| Help you to relax (RELAX) |
96.8i |
13.1 |
29.9 |
17.2 |
28.6 |
82.7ii |
| Help you feel more confident or more able to talk to people in a social situation (INCREASE CONFIDENCE) |
36.5 |
53.1 |
42.0 |
10.3 |
66.1 |
70.2v |
| Improve the effects of other substances (IMPROVE EFFECTS) |
44.3 |
37.5 |
27.4 |
29.3 |
26.2 |
41.0 |
| Help ease the after effects of other substances (AFTER EFFECTS) |
64.6 |
11.9 |
8.3 |
3.4 |
12.5 |
35.9 |
| Help you to stay awake (STAY AWAKE) |
7.5 |
91.3ii |
72.0iv |
50.0 |
69.0ii |
10.6 |
| Help you lose your inhibitions (LOSE INHIBITIONS) |
28.1 |
41.9 |
49.7 |
36.2 |
41.1 |
51.0 |
| Enhance feelings when having sex (ENHANCE SEX) |
27.8 |
31.9 |
63.1 |
25.9 |
52.4 |
32.1 |
| Help you stop worrying about a problem (STOP WORRYING) |
57.7 |
22.5 |
32.5 |
15.5 |
41.7 |
55.1 |
| Help make something you were doing less boring (DECREASE BOREDOM) |
70.1v |
45.6 |
36.3 |
44.8 |
53.6 |
60.6 |
| Help you to sleep (SLEEP) |
69.6iv |
– |
– |
– |
– |
30.4 |
| Help you to concentrate or to work or study (WORK) |
20.9 |
29.4 |
3.2 |
1.7 |
6.0 |
6.4 |
| Enhance an activity such as listening to music or playing a game or sport (ENHANCE ACTIVITY) |
72.8iii |
66.2iii |
79.6ii |
72.4iii |
60.7 |
51.6 |
| Total number of items in the scale |
17 |
17 |
17 |
16 |
17 |
17 |
| Chronbach’s α for scale items |
0.78 |
0.74 |
0.76 |
0.73 |
0.78 |
0.84 |
| Mean total number of different functions endorsed for use of [substance] (range) |
9.0 (0–17) |
7.8 (0–16) |
8.0 (0–16) |
5.9 (0–15) |
7.9 (0–17) |
8.8 (0–17) |
Braakman M, Kortmann F, van den Brink W. 2009. “Validity of ‘post-traumatic stress disorder with secondary psychotic features’: a review of the evidence” Acta Psychiatr Scand 119(1): 15-24
http://www.ncbi.nlm.nih.gov/pubmed/18764840/
Twenty-four comparative studies were included. These studies indicate that PTSD-SP is a syndrome that comprises PTSD-symptoms followed in time by the additional appearance of psychotic features. The psychotic features are not confined to episodes of re-experiencing, but remain present continuously. PTSD-SP seems to have some biological features differentiating it from schizophrenia and PTSD, e.g. there are differences in smooth pursuit eye movement patterns, concentrations of corticotropin-releasing factor and dopamine beta-hydroxylase activity
Bradley A, Dinan T. 2010. “Review: A systematic review of hypothalamic-pituitary-adrenal axis function in schizophrenia: implications for mortality” Journal of Psychopharmacology 24: 91
http://www.ncbi.nlm.nih.gov/pubmed/20923924 in dropbox
Brandt C, Eichele T, Melle I, Sundet K, Server A, Agartz I, Hugdahl K, Jensen J, Andreassen O. 2014. “Working memory networks and activation patterns in schizophrenia and bipolar disorder: comparison with healthy controls” British Journal of Psychiatry 204: 290-298
http://bjp.rcpsych.org/content/204/4/290.long in dropbox
patients with schizophrenia (n = 100) and bipolar disorder (n = 100) and a healthy control group (n = 100) performed a 2-back working memory task while fMRI data were acquired
Similar working memory networks were activated in all groups. However, in three out of nine networks related to the experimental task there was a graded response difference in fMRI signal amplitudes, where patients with schizophrenia showed greater activation than those with bipolar disorder, who in turn showed more activation than healthy controls
In both disorders the impairment is stable over time and across different clinical states, is relatively resistant to treatment and contributes to poor functional outcome.5
dorsolateral (Brodmann areas (BAs) 9/46) and ventrolateral (BAs 44/45/47) prefrontal cortex, premotor cortex (BAs 6/32) and parietal cortex (BAs 7/40) in both hemispheres.15,16 The last three components were deactivations in regions consistent with the main sites of the default mode network.29
no disease-specific network or compensatory region was detected
brain activation of people with schizophrenia and healthy controls may be represented by different, but overlapping, inverted U-curve shapes of activation.31,32 According to such a model, patients will show stronger activation than controls even if task difficulty is low
The three components that showed group differences were all in frontal and parietal brain regions consistent with the well-known working memory network
Breetvelt E, Boks M, Numans M, Selten J, Sommer I, Grobbee D, Kahn R, Geerlings M. 2010. “Schizophrenia risk factors constitute general risk factors for psychiatric symptoms in the population” Schizophr Res 120(1-3): 184-90
http://www.ncbi.nlm.nih.gov/pubmed/20421160
general population sample of 4894 subjects (mean age 39 years, 45% men) from the Utrecht Health Project
Participants with non-clinical psychotic symptoms had an 89% probability of concomitant depressive, anxiety or phobic anxiety symptoms, compared to 11% in participants without psychotic symptoms. The risk profiles for non-clinical psychotic symptoms and other psychiatric symptoms were largely similar. Non-Dutch ethnicity was most strongly associated with non-clinical psychotic symptoms. Adjusting for other psychiatric symptoms did not increase the specificity of the risk factors
Socio-demographic risk factors for non-clinical psychotic symptoms in the general population are also risk factors for other psychiatric symptoms
Bremner J, Southwick S, Darnell A, Charney D. 1996. “Chronic PTSD in Vietnam combat veterans: course of illness and substance abuse” Am J Psychiatry 153(3): 369-75
http://www.ncbi.nlm.nih.gov/pubmed/8610824
substance use increased in parallel with PTSD symptoms; alcohol, marijuana, heroin, and benzodiazepines reported to make PTSD symptoms better, while cocaine made symptoms in the hyperarousal category worse
Bremner J, Vythilingam M, Vermetten E, Southwick S, McGlashan T, Nazeer A, Khan S, Vaccarino L, Soufer R, Garg P, Ng C, Staib L, Duncan J, Charney D. 2003. “MRI and PET study of deficits in hippocampal structure and function in women with childhood sexual abuse and posttraumatic stress disorder” Am J Psychiatry 160 (5): 924-32
http://ajp.psychiatryonline.org/article.aspx?articleid=176214
women with sexual abuse and PTSD (N=10) 16% smaller hippocampus than PTSD- abuse+ (N=12), 19% smaller than abuse- (N=11)
Breslau N, Davis G, Schultz L. 2003. “Posttraumatic stress disorder and the incidence of nicotine, alcohol and other drug disorders in persons who have experienced trauma” Arch Gen Psychiatry 60(3): 289-294
http://archpsyc.jamanetwork.com/article.aspx?articleid=207281
The prospective and retrospective data show an increased risk for the onset of nicotine dependence and drug abuse or dependence in persons with PTSD, but no increased risk or a significantly (P=.004) lower risk (for nicotine dependence, in the prospective data) in persons exposed to trauma in the absence of PTSD, compared with unexposed persons
Breslau N, Anthony J. 2007. “Gender differences in the sensitivity to posttraumatic stress disorder: an epidemiological study of urban young adults” Journal of Abnormal Psychology 116(3): 607-611
http://www.epi.msu.edu/janthony/K05/Breslau_Gender%20differences%20in%20the%20sensitivity%20ptsd.pdf
Qualifying events occur more often among men than among women, though the gender gap (i.e., male excess) tends to be small except for assaultive violence (e.g., shot/stabbed, badly beaten up, mugged/threatened with a weapon, raped), which occurs far more often among men (Breslau, Kessler, Chilcoat, et al., 1998; Breslau, Wilcox, et al., 2004)
Of the total sample of 1,698 young adults, 82.5% (n 1,401; 87.2% of men and 78.4% of women) were exposed to one or more DSM–IV qualifying traumatic events. The mean number of traumatic events was 4.8 (6.1 per male respondent and 3.7 per female respondent)
Among men, the conditional probability of PTSD varied little by (a) whether the worst event involved assaultive violence (7.2% vs. 7.7%)
among women, the conditional probability of PTSD following an assaultive violence event was markedly higher than following a nonassaultive event, 24.3% versus 6.9%
Britton J, Phan K, Taylor S, Fig L, Liberzon I. 2005. “Corticolimbic blood flow in posttraumatic stress disorder during script-driven imagery.” Biol Psychiatry. 57(8): 832-40
http://www.ncbi.nlm.nih.gov/pubmed/15820703/
PTSD patients had greater rostral anterior cingulate (rACC) deactivation
Brodbeck J, Matter M, Page J, Moggi F. 2007. “Motives for cannabis use as a moderator variable of distress among young adults.” Addict Behav 32(8): 1537-45
http://www.ncbi.nlm.nih.gov/pubmed/17178197
2031 young Swiss adults; only cannabis users with coping motives showed lower mental health, greater pathology, more distress, and more life events than non-users whereas those with predominantly social reasons for use were not more distressed than non-users
Our results suggest that secondary prevention for cannabis users should target especially young adults with coping motives for use
Brook D, Brook J, Zhang C, Cohen P, Whiteman M. 2002. “Drug use and the risk of major depressive disorder, alcohol dependence and substance use disorders” Arch Gen Psychiatry 59(11): 1039-1044
http://archpsyc.jamanetwork.com/article.aspx?articleid=206845
community-based sample of 736 adults (50% female) from upstate New York, the subjects were interviewed at the mean ages of 14, 16, 22, and 27 years
Adolescent and young adult tobacco use was significantly associated with an increased risk of alcohol dependence and substance use disorders at a mean age of 27 years, but not with new episodes of major depressive disorder. Earlier alcohol use significantly predicted later major depressive disorder, alcohol dependence, and substance use disorders in the late 20s, as did early marijuana use and other illicit drug use. Except for the effect of tobacco use on major depressive disorder, early drug use was significantly related to later psychiatric disorders, even after statistically controlling for age, sex, parental educational level, family income, and prior episodes of major depressive disorder and substance use disorders
Marijuana use during childhood and early adolescence (OR, 1.18; 95% CI, 1.04-1.33; P<.05), middle and late adolescence (OR, 1.23; 95% CI, 1.06-1.43; P<.05), and the early 20s (OR, 1.41; 95% CI, 1.21-1.65; P<.001) also showed substantial effects on later MDD, alcohol dependence, and SUDs
the study provides evidence that drug use precedes MDD and alcohol dependence, and is not merely a consequence of these disorders
the similarity of the ORs between drug use and psychiatric disorders (1.2-1.4) suggests the homogeneity of their mechanisms, which may arise from unmeasured common underlying factors
a decrease in youth substance use may be accompanied by a corresponding decrease in later psychiatric disorders
Brown G, Craig T, Harris T, Handley R, Harvey A. 2007. “Development of a retrospective interview measure of parental maltreatment using the Childhood Experience of Care and Abuse (CECA) instrument — a life-course study of adult chronic depression -1” J Affect Disord 103(1-3): 205-15
http://www.ncbi.nlm.nih.gov/pubmed/17651811/
adverse maternal behaviour emerges as of critical importance for the link with adult chronic depression. Maternal lack of affection (‘neglect’) and maternal rejection (’emotional abuse’) form the core of an index of parental maltreatment, and it is concluded that persistent rejection, particularly from a mother, appears to be the core experience of importance
Broyd S, Demanuele C, Debener S, Helps S, James C, Sonuga-Barke E. 2009. “Default-mode brain dysfunction in mental disorders: a systematic review” Neuroscience and Biobehavioral Reviews 33: 279-296
http://www.ncbi.nlm.nih.gov/pubmed/18824195 in dropbox
Marcus Raichle first coined the term ‘default-mode’ in relation to resting state brain function ( Raichle et al., 2001 )
attenuation of the ventral MPFC occurred with tasks involving judgments that were self-referential, while activity in the dorsal MPFC increased for self-referential stimuli, suggesting the dorsal MPFC is associated with introspective orientated thought ( Gusnard et al., 2001).
the DMN is characterised by very low frequency neuronal oscillations providing temporal synchrony between functionally specific and diverse brain regions (Sonuga-Barke and Castellanos, 2007)
Should only the task-negative network be termed the DMN and contrasted with the task-positive network? Or should both task-positive and task-negative networks be regarded as elements of the DMN? … we use the DMN term to describe the task-negative network specifically. We use the term Low Frequency Resting State Networks (LFRSN) to describe both the task-positive and task-negative networks.
The human brain then, is never really ‘at rest’, but rather an orchestra of distinct functional networks in dynamic concert
PCC appears to serve an important adaptive function and is implicated in broad-based continuous sampling of external and internal environments ( Raichle et al., 2001), the attenuation of which during transition from rest facilitates focused attention during task-specific activity (Eichele et al., 2008; Raichle et al., 2001). Moreover, modulation of the PCC during working-memory tasks (Greicius et al., 2003 ), reduced activation at rest (Greicius et al., 2004 ), susceptibility to atrophy in Alzheimer’s disease patients (Buckner et al., 2005), and reduced connectivity with anterior DMN regions in attention deficit/ hyperactivity disorder (ADHD) participants ( Castellanos et al., 2008; Uddin et al., 2008a ) suggests that this region may be implicated in working memory dysfunction. Finally, PCC and retrosplenial cortex are also associated with the processing of emotionally salient stimuli, and may play a role in emotional processing related to episodic memory (Maddock, 1999)
MPFC is thought to mediate a dynamic interplay between emotional processing and cognition functions which map on to the ventral and dorsal regions, respectively ( Gusnard et al., 2001; Raichle et al., 2001; Simpson et al., 2001). Ventral MPFC is heavily interconnected with other limbic structures such as amgydala, ventral striatum and hypothalamus, which Gusnard and colleagues suggest may indicate a role in relation to the mediation of visceromotor aspects of emotional information gathered from external and internal sources ( Gusnard et al., 2001; Gusnard and Raichle, 2001). Both PCC/precuneus and MPFC are associated with introspective processes such as self-referential and emotional processing, and are attenuated when attention is directed toward external events ( Gusnard and Raichle, 2001)
In view of the lack of association between alpha and resting state brain activity, Laufs et al. (2003b) hypothesise that alpha may act as a baseline for specific brain structures associated with the
attentional system, and more specifically the task-positive net-work ( Laufs et al., 2006 ). In contrast, Mantini et al. (2007) reported positive correlations between beta (13–30 Hz) and alpha (8–13 Hz) with the PCC, precuneus, bilateral superior frontal gyrus and the MFG
The results ofMeltzer et al. (2007) and Scheeringa et al. (2008) suggest that the DMN may operate within the theta frequency such that decreased DMN activation is associated with increased theta power during goal-directed activity
low frequency electromagnetic activity is notoriously difficult to study. Interference to the low frequency fluctuations in the BOLD signal may arise from low frequency cycles in respiration and cardiac activity, and the related body and head movements may obscure EEG and MEG signals. Variations in respiration ( 0.03 Hz) correlate highly with fluctuations in the DMN ( <0.1 Hz), with overlap between regions within the DMN and areas significantly affected by changes in respiration (Birn et al., 2006, 2008)
The ability to maintain attentional focus and resist distraction or lapses of attention is conventionally considered to underlie higher order top–down control… two groups have suggested that the DMN may represent a mechanism underlying deficient top–down attentional control (Mason et al., 2007; Sonuga-Barke and Castellanos, 2007). Both perspectives predict that attentional lapses during goal-directed action may be a result of interference arising from spontaneous, and most likely self-referential, thought
Spontaneous low frequency activity in the task-negative compo-nent of the default-mode network which is routinely attenuated during goal directed tasks, can under certain circumstances (e.g. suboptimal motivational states or in individuals with attention disorders) persist into or remerge during periods of task-related active processing to such an extent that it competes with task-specific neural processing and creates the context for periodicattentional intrusions/lapses and cyclical deficits in performance; the temporal signatures of task-negative fluctuations being mirrored in patterns of attention and performance ( Sonuga-Barke and Castellanos, 2007, p. 981)
lapses in attention will be caused by intrusions of introspective thought which produce increased variability in task performance
The notion of wandering minds provides another possible theory of attentional lapses during task performance, and it is conceivable that generalised deficits in attention and cognitive control may well coincide with an increased incidence of stimulus-independent thought or mind wandering and the intrusion of increased activity in the DMN
In a group of schizophrenic patients, Zhou et al. (2007) reported significantly increased resting state connectivity in the task-positive network and DMN in the patient group, alongside significantly increased anti-correlations between the networks. These results were more specifically associated with bilateral dorsal MPFC, inferior temporal gyrus, and lateral parietal region in the DMN; and right dorsal lateral PFC and bilateral insula and orbital frontal gyrus in the task-positive network. The authors argue that the increased anti-correlations reflect increased antagonism and excessive competition between the networks, and most likely contribute to the over-mentalising and deficit in attentional control symptomatic of schizophrenia. More interest-ingly, reduced anti-correlations were evident between the right dorsal premotor cortex and the PCC and bilateral parietal region of the DMN and suggest that the right dorsal premotor cortex may be instructive in mediating the anti-correlation between the DMN and task-positive networks (Zhou et al., 2007 )
resting state BOLD fMRI scans revealed a reduced anti-correlation in an ADHD population relative to controls, thought to relate to attentional lapses and performance variability sympto-matic of this disorder (Castellanos et al., 2008)
Kennedy and Courchesne (2008) report no significant anti-correlations in a sample of individuals with autism spectrum disorder (ASD). This patient group did not show any abnormal patterns of connectivity in the task-positive network, but did show reduced connectivity in the DMN mainly associated with the MPFC
Hahn et al. (2007) reported nicotine to increase deactivation within the DMN for task-specific cues, indicating increased preparation and improvements in attentional control ( Hahn et al., 2007)
excessive rivalry between the functions subserved by the networks and associated neural processing, will be accompanied by cyclical fluctuations in attention and deficits in goal-directed action. Generally, this will manifest as an attentional deficit
In Alzheimer’s disease, connectivity between the right hippocampus and many component regions of the DMN, including the dorsal MPFC, ventral ACC, middle temporal gyrus (MTG) and the right PCC is reduced ( Wang et al., 2006), and likely relates to a dysfunction of working memory and attentional processes. Similarly, although one study has reported increased resting state functional connectivity within the DMN in adoles-cents with ADHD (Tian et al., 2006 ), more commonly, aberrant functional connectivity between the anterior (e.g. MFPC and superior frontal gyrus) and posterior (e.g. PCC/precuneus) compo-nents of the DMN is observed ( Castellanos et al., 2008; Uddin et al., 2008a )
in schizophrenia, increased connectivity between the DMN and other resting state networks suggests greater depen-dence on other resting state networks, most likely associated with increased distraction due to hallucinations and delusional experiences (Jafri et al., 2008 ). Moreover, increased connectivity within the DMN and task-positive network in schizophrenic patients at rest implies that these individuals are susceptible to over-mentalising and excessive alertness to the external environ-ment, respectively (Zhou et al., 2007 )
aberrant increases in connectivity will likely manifest in delusional experience, over ardent mentalising, and attentional lapses due to the disproportionate involvement and interaction between the DMN and associated networks
n schizophrenia, positive symp-tom severity was correlated with increased deactivation in the MFG, precuneus, and the left MTG in an oddball task (Garrity et al., 2007), while impaired self-monitoring processes and stimulus-independent thought have been associated with abnormal low frequency resting state connectivity (Bluhm et al., 2007; Liang et al., 2006 ). In autism however, atypical or reduced self-referential, affective and introspective thought is associated with low activation of the DMN in the resting state. Additionally, atypical emotional processing in autism is notable by the absence of any functional activation of the medial orbital frontal component of the DMN during the processing of emotional compared with neutral words in a Stroop task (Kennedy et al., 2006). Indeed, reduced task-induced deactivation of the DMN during a number-word Stroop task has been associated with greater social impairment in participants with autism (Kennedy et al., 2006 ). More specifically, reduced DMN connectivity in autism is associated with the MPFC
In depressed patients, the subgenal cingulate is a prominent region within the DMN and related to the length of depressive episode, while noticeably absent in the DMN of control partici-pants (Greicius et al., 2007 ). Further, in this patient group increased connectivity between the DMN and the thalamus is argued to reflect increased interaction between the thalamus and subgenal cingulate due to increased emotional processing, at the expensive of executive functions (Greicius et al., 2007 ). In patients with anxiety disorders, reduced deactivation of MPFC and increased deactivation of the PCC is thought to be indicative of increased levels of anxiety (MPFC) and emotional processing (PCC) of task-specific threat words
reduced connectivity between MPFC and PCC regions of the DMN is associated with aging, and the anterior region of the DMN is correlated with cognitive decline. In Alzheimer’s patients, the hippocampus features as a prominent node within the DMN and shows reduced connectivity with other DMN regions
greater deactivation in the MFG, precuneus, and the left MTG were correlated with positive symptoms of schizophrenia (Garrity et al., 2007 )
increased connectivity between the DMN and other resting state networks using spatial ICA, has been hypothesised to reflect distraction and hallucinatory experiences of schizophrenic patients (Jafri et al., 2008). … reduced deactivation of ACC was suggested to be associated with reduced attentional control
subgenual cingulate was found to contribute disproportionally to the connectivity of the DMN in this patient group, with increases in connectivity associated with depression refractoriness, or the length of the current depressive episode (Greicius et al., 2007). There was also increased functional connectivity in the thalamus during rest. Greicius et al. (2007) suggest that increased
connectivity in ‘affective’ regions may detrimentally affect connectivity in regions associated with cognitive processing such as the dorsal anterior cingulate (Greicius et al., 2007)
no difference in the deactivation of the DMN was evident between a group of anxiety patients or controls when emotionally neutral words were alternated with rest. In contrast, when threat words were alternated with emotionally neutral words, anxiety patients demonstrated reduced deactiva-tion of the MPFC and increased deactivation of the PCC relative to controls ( Zhao et al., 2007)
it has been postulated that a deficit in the mirror neuron network is associated with DMN dysregulation in autism, suggesting that this disorder may arise from an atypical processing of self and relationship with others (Iacoboni, 2006 )
increased functional connectivity with dorsal ACC was observed for bilateral dorsal ACC, bilateral thalamus, bilatertal cerebellum, bilateral insula and bilateral brain stem in the ADHD compared to control group… the ADHD group showed less negatively correlated functional activity in the ACC, MFG and superior temporal gyrus… may indicate a relationship between working
memory deficits and attentional lapses in ADHD (Castellanos et al., 2008)… adult ADHD and control groups did not differ on global measures of DMN homogeneity, although significantly reduced network homogene-ity in the posterior regions of the DMN, particularly the precuneus was observed in the ADHD group (Uddin et al., 2008a)
Burstein S, Hunter S. 1995. “Stimulation of anandamide biosynthesis in N-18TG2 neuroblastoma cells by delta 9-tetrahydrocannabinol (THC)” Biochem Pharmacol 49(6): 855-8
http://www.ncbi.nlm.nih.gov/pubmed/7702643
upregulation
Butters J. 2002. “Family stressors and adolescent cannabis use: a pathway to problem use” J Adolesc 25(6): 645-54
http://www.ncbi.nlm.nih.gov/pubmed/12490182
those reporting family disruption (e.g.,from death or divorce) 79% more likely to use cannabis; of those who were already using cannabis, … family disruption were 87% more likely to progress to problem use.
van Buuren M, Gladwin T, Zandbelt B, van den Huevel M, Ramsey N, Kahn R, Vink M. 2009. “Cardiorespiratory effects on default-mode network activity as measured with fMRI” Human Brain Mapping 30: 3031-3042
http://www.ncbi.nlm.nih.gov/pubmed/19180557 in dropbox
The default-mode network (DMN) consists of areas showing more activation during rest than during a task
The goal of this study is to investigate whether BOLD signal changes within the DMN can be explained by CR effects
CR effects were present within the frequency-range of the DMN and were located in areas of the DMN, but equally so in other areas. After removal of CR effects, deactivation and resting-state connectivity between the areas of the DMN remained significant. In addi-tion, DMN deactivation was still modulated by task demand. The same CR correction method did remove activation in task-related areas. We take these results to indicate that the BOLD signal within the DMN cannot be explained by CR effects alone and is possibly related to some form of cognitive neuronal processing
Functional imaging studies typically investigate brain function by identifying areas showing increased activation during a task as compared to rest or baseline. However, some areas in the brain are more active during rest [Binder et al., 1999; Mazoyer et al., 2001; Shulman et al., 1997]. To-gether, these areas are called the default-mode network (DMN) and include the posterior cingulate (PCC), the medial prefrontal cortices and the lateral posterior cortices (Brodmann area 39) [Fox et al., 2005; Greicius et al., 2003; Raichle et al., 2001]
the cardiac and respiratory cycle cause signal changes in brain areas near major vessels and the ventricles, and motion artifacts near the edges of the brain [Bhattacharyya and Lowe, 2004; Dagli et al., 1999; Glover et al., 2000; Hu et al., 1995]. In addition, fluctuations in heart rate (HR), heart rate variability (HRV) and respiration volume are related to BOLD signal changes in grey matter areas not located near large vessels [Birn et al., 2006; Critchley et al., 2003; Shmueli et al., 2007; Wise et al., 2004]
correcting for CR effects resulted in both reduced BOLD signal changes during the task (Fig. 5A,B) and reduced resting-state connectivity in the DMN and the task-positive network (Table III) (i.e. areas that showed more activation during the task compared to rest). However, even after applying a strict method to cor-rect for CR effects, BOLD signal changes within the DMN
during the task as well as resting-state connectivity between the areas of the DMN remained significant. In addition, DMN deactivation was still modulated by the amount of effort or attention required to perform the task
the proportion of variance explained by factors modeling CR effects was higher within the task-positive network compared to DMN
BOLD signal changes within the DMN were correlated with EEG power within the alpha and beta band [Mantini et al., 2007]
This study confirmed that part of the BOLD signal within the DMN originates from confounding effects of cardiorespiratory processes
Cabral G. 2001. “Marijuana and Cannabinoids: Effects on Infections, Immunity, and AIDS” Journal of Cannabis Therapeutics 1(3/4): 61-85
Cabral G, Raborn E, Griffin L, Dennis J, Marciano-Cabral F. 2008. “CB2 receptors in the brain: role in central immune function” Br J Pharmacol 153(2): 240-251
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2219530/
Recently, it has been recognized that the cannabinoid receptor CB2 may play a functionally relevant role in the central nervous system (CNS). This role is mediated primarily through microglia, a resident population of cells in the CNS that is morphologically, phenotypically, and functionally related to macrophages. These cells also express the cannabinoid receptor CB1. The CB1 receptor (CB1R) is constitutively expressed at low levels while the CB2 receptor (CB2R) is expressed at higher levels and is modulated in relation to cell activation state. The relatively high levels of the CB2R correspond with microglia being in ‘responsive’ and ‘primed’ states, suggesting the existence of a ‘window’ of functional relevance during which activation of the CB2R modulates microglial activities. Signature activities of ‘responsive’ and ‘primed’ microglia are chemotaxis and antigen processing, respectively. The endocannabinoid 2-arachidonylglycerol has been reported to stimulate a chemotactic response from these cells through the CB2R. In contrast, we have shown in vivo and in vitro that the exogenous cannabinoids delta-9-tetrahydrocannabinol and CP55940 inhibit the chemotactic response of microglia to Acanthamoeba culbertsoni, an opportunistic pathogen that is the causative agent of Granulomatous Amoebic Encephalitis, through activation of the CB2R
CB1R is G-protein coupled as evidenced by inhibition of adenylyl cyclase (Howlett et al., 1986), inhibition of N-type calcium channels (Mackie and Hille, 1992) and increased binding of non-hydrolyzable GTPγS in the presence of cannabinoids (Sim et al., 1996)
Transcripts (that is, mRNAs) for the CB1R and CB2R have been found in spleen and tonsils (Munro et al., 1993; Galiègue et al., 1995) and other immune tissues (Munro et al., 1993; Bouaboula et al., 1996). However, in all cases reported to date, levels of message for the CB2R in immune cells exceed those for the CB1R. The distribution pattern of levels of CB2R mRNA displays major variation in human blood cell populations with a rank order of B lymphocytes>natural killer cells≫monocytes>polymorphonuclear neutrophils>CD8 lymphocytes>CD4 lymphocytes (Galiègue et al., 1995)
The presence of the CB2R primarily within immune cells suggests a role for this receptor in the activities attributed to these cells. However, CB1R also may be involved in cannabinoid-mediated modulation of select immune functions (Stefano et al., 1996; Sinha et al., 1998; Waksman et al., 1999)
palmitoylethanolamide, an analogue of anandamide that does not bind the CB2R, causes a reduction of pain associated with inflammatory response (Calignano et al., 1998; Jarai et al., 1999) that is blocked by the CB2R antagonist ((1S-endo)-5-(4-Chloro-3-methylphenyl)-1-((4-methylphenyl)methyl)-N-(1,3,3-trimethylbicyclo(2.2.1)hept-2-yl)-1H-pyrazole-3-carboxamide (SR144528). The nuclear peroxisome proliferator-activated receptor-alpha (PPAR-α) has been reported as the mediator of the anti-inflammatory actions of this lipid amide (Lo Verme et al., 2005a, 2005b)
Microglia migrate and proliferate during and after injury and inflammation (Leong and Ling, 1992; Kreutzberg, 1995, 1996; Benveniste, 1997a). Once activated, they produce various cytokines including interleukin (IL)-1, IL-6 and tumour necrosis factor-α (Giulian et al., 1986; Reid et al., 1993; Benveniste, 1997b), and express major histocompatability complex (MHC) class I and II antigens and the complement receptor, CD11/CD18 complex. Microglia, also, are phagocytic and, upon activation, can process antigens and exert cytolytic functions. Paradoxically, these cells not only play a role in host defense and tissue repair in the CNS (Streit et al., 1988; Perry, 1990), but also have been implicated in nervous system disorders, such as Multiple Sclerosis (MS) (Matsumoto et al., 1992), Alzheimer’s disease (Rogers et al., 1988), Parkinson’s disease (McGeer et al., 1988) and acquired immune deficiency syndrome (AIDS) dementia (Dickson et al., 1991; Merrill and Chen, 1991; Spencer and Price, 1992). Neural histological features of AIDS dementia complex include diffuse leukoencephalopathy of the white matter, which is accompanied by severe loss of myelin and sparing of fibres (Kleihues et al., 1985) Discrete areas of demyelination with hypertrophied astrocytes, which also contain microglia, blood-derived macrophages and multinucleated giant cells, are observed. The multinucleated giant cells have been reported to constitute syncytia of macrophages or microglia, which are productively infected with the human immunodeficiency virus type 1 (HIV1) and are a histopathological hallmark of subacute encephalitis in HIV1-infected brains (Koenig et al., 1986; Michaels et al., 1988) and spinal cords (Eilbott et al., 1989; Maier et al., 1989). Thus, it has been proposed that in AIDS dementia, macrophages and microglia are the predominant cell types, which are infected and produce HIV1 (Koyanagi et al., 1987; Kure et al., 1990)
The CB2R is not detected in ‘resting’ cells, is present at high levels in ‘responsive’ and ‘primed’ cells, and is identified at greatly diminished levels in ‘fully’ activated cells. In contrast, the CB1R is present in microglia at relatively low levels and is expressed constitutively in relation to cell activation state
since the kinetics of CB1R and CB2R expression by microglia are distinctive, activation of the two receptors by endogenous and/or exogenous cannabinoids may result in disparate functional outcomes
fully’ activated microglia are not susceptible to cannabinoid-mediated action that is linked to the CB2R
Waksman et al. (1999) reported that the production of inducible nitric oxide (iNO), a potent inflammatory mediator that is released from microglia and macrophage-like cells upon their ‘full’ activation, was inhibited by cannabinoids in a mode that was linked, at least in part, to the CB1R
Puffenbarger et al. (2000) extended these studies on the effects of cannabinoids on ‘fully’ activated microglia and indicated that the inhibition of the inducible expression of pro-inflammatory cytokines was exerted through a non-CB1, non-CB2 receptor process
Exposure of neonatal rat cortical microglia to THC resulted in reduced amounts of lipopolysaccharide-induced mRNAs for IL-1α, IL-1β, IL-6 and tumour necrosis factor-α. Of these cytokine mRNAs, the response of that for IL-6 was exquisitely sensitive to THC treatment… when methanandamide, the non-hydrolyzable analogue of anandamide, was tested, its ability to inhibit cytokine mRNA expression was comparable to that of THC
Stefano et al. (1998) reported that acute exposure to anandamide resulted in transformation of macrophages from an amoeboid and motile state to that of a rounded and non-motile conformation. These investigators proposed that the transforming events were linked to the CB1R since the CB1R-selective antagonist SR141716A blocked the transformation
The chemotactic response of mouse macrophages to formal-methionyl-leucine-phenylalanine also has been shown to be decreased by CBD (Sacerdote et al., 2005), a cannabinoid that binds weakly to the CB2R. The CB2R antagonist SR144528 prevented this decrease, suggesting a functional linkage to the cognate receptor
Walter et al. (2003) found that the endocannabinoid 2-arachidonylglycerol (2-AG) triggered migration of microglia and that the CB2R was involved in this effect
Franklin and Stella (2003) suggested that CB2Rs and ‘abnormal-CBD-sensitive’ receptors regulated the migration of microglial-like cells. Stella and co-workers extended these studies and showed that P2X7 ionotropic receptors played a key role in controlling the production of 2-AG by microglia (Witting et al., 2004). Recently, Raborn et al. (in press) demonstrated that THC and CP55940 mediated inhibition of mouse peritoneal macrophage chemotaxis to the chemokine RANTES/CCL5 and that this event was linked to the CB2R.
cannabinoids act through the CB2R to alter macrophage migration with exogenous cannabinoids, such as THC exerting inhibitory effects and endocannabinoids such as 2-AG eliciting an opposite stimulatory effect
the CB2R may be a constituent element of a network of G-protein-coupled receptor signal transductional systems, inclusive of chemokine receptors, that act coordinately to modulate macrophage migration
THC impaired the ability of a macrophage hybridoma to function as an antigen-presenting cell based on its ability to secrete IL-2 upon stimulation of a soluble protein antigen-specific helper T-cell hybridoma… CB2 activation on macrophages
CB2R as playing a relevant functional role in the early inflammatory process by macrophages and macrophage-like cells, namely chemotaxis and antigen-processing functional attributes of these cell types when in ‘responsive’ and ‘primed’ states. Since microglia constitute a resident population of macrophages in the brain, exhibit phenotypic and functional properties of macrophages, and express the CB2R at maximal levels when in ‘responsive’ and ‘primed’ states, a ‘window’ of functional relevance for the CB2R comparable to that for macrophages at peripheral sites may be operative
Mice treated with THC exhibited higher mortalities from infection with Acanthamoebaas compared to similarly infected vehicle control mice (Marciano-Cabral et al., 2001)
The CB2R, but not the CB1R, agonist inhibits chemotaxis of microglia
It is postulated that bioactive lipid mediators thus generated include the endocannabinoid 2-AG that serves to drive chemotaxis by autocrine and/or paracrine activation of the CB2R. The exogenous cannabinoid THC may inhibit this chemotactic response by superimposing a signal transductional activation of the CB2R. That is, THC could inhibit the synthesis and/or release of 2-AG or, alternatively, by virtue of its relative long half-life as compared to that of 2-AG, preclude this endocannabinoid from ligating to the CB2R
Alzheimer’s disease, Parkinson’s Disease, MS, amyotrophic lateral sclerosis and HIV-associated dementia. The pathological hallmark of these diseases is chronic inflammation induced by persistent cell activation and elicitation of proinflammatory mediators (that is, NO, cytokines and chemokines). Studies have shown that microglia and microglia-derived cells are the major cell types responsible for this neuroinflammation
During activation, microglia upregulate an array of cell-surface receptors that may be critical in microglial regeneration and/or degeneration of the CNS. Included among these are immunoglobulin (Ig) superfamily receptors, complement receptors, toll-like receptors, cytokine/chemokine receptors, opioid receptors and cannabinoid receptors. Microglia have been found to express both the CB1R and CB2R in vitro (Carlisle and Cabral, 2002; Carrier et al., 2005) and to produce the endocannabinoids 2-AG as well as anandamide in lesser quantities (Carrier et al., 2004)… Activation of the CB2R on these cells appears to promote migration and proliferation. Walter et al. (2003) demonstrated that 2-AG induced migration of microglia and that this occurred through the CB2R and abnormal-CBD-sensitive receptors
Fernandez-Ruiz et al. (2007), using a variety of neurodegenerative disease models, reported the expression of the CB2R in microglia, astrocytes and neuron sub-populations. This expression of the CB2R in vivo apparently is attributed, in large measure, to microglia. In several neurodegenerative diseases, upregulation of microglial CB2R has been observed (Zhang et al., 2003; Benito et al., 2005, 2007; Maresz et al., 2005; Yiangou et al., 2006; Ashton et al., 2007)
It has been proposed that the role of the CB2R in immunity in the CNS is primarily that of anti-inflammatory (Carrier et al., 2005)
Cabral G, Griffin-Thomas L. 2009. “Emerging role of the CB2 cannabinoid receptor in immune regulation and therapeutic prostpects” Expert Rev Mol Med 11: e3
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2768535/
CB2 may serve as a selective molecular target for therapeutic manipulation of untoward immune responses including those associated with a variety of neuropathies that exhibit a hyperinflammatory component
Campolongo P, Roozendaal B, Trezza V, Hauer D, Schelling G, McGaugh J, Cuomo V. 2009. “Endocannabinoids in the rat basolateral amygdala enhance memory consolidation and enable glucocorticoid modulation of memory.” Proc Natl Acad Sci U S A 106(12): 4888-93
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2660732/
basolateral amygdala modulates consolidation of emotionally arousing memories, involving glucocorticoids. CB1 agonists enhance, and antagonists impair, retention of avoidance training if injected into rat BLA after training. Antagonist blocks corticosterone memory enhancement
Campos A, Ferreira F, Guimaraes F, Lemos J. 2010. “Facilitation of endocannabinoid effects in the ventral hippocampus modulates anxiety-like behaviors depending on previous stress experience” Neuroscience 167(2): 238-46
http://www.ncbi.nlm.nih.gov/pubmed/20167262/
AM404 (5-50 pmol) microinjection promoted an anxiogenic-like effect in non-stressed rats but decreased anxiety in stressed animals
Cardno A, Gottesman I. 2000. “Twin studies of schizophrenia: from bow-and-arrow concordances to Star Wars Mx and functional genomics” American Journal of Medical Genetics 97 (1): 12-17
http://onlinelibrary.wiley.com/doi/10.1002/(SICI)1096-8628(200021)97:1%3C12::AID-AJMG3%3E3.0.CO;2-U/abstract
concordance rates of 41–65% in monozygotic (MZ) pairs and 0–28% in dizygotic (DZ) pairs, and heritability estimates of approximately 80–85%
Carey C, Agrawal A, Zhang B, Conley E, Degenhardt L, Heath A, Li D, Lynskey M, Martin N, Montgomery G, Wang T, Bierut L, Hariri A, Nelson E, Bogdan R. 2015. “Monoacylglycerol lipase (MGLL) polymorphism rs604300 interacts with childhood adversity to predict cannabis dependence symptoms and amygdala habituation: evidence from an endocannabinoid system-level analysis” J Abnorm Psychol 124(4): 860-77
http://www.ncbi.nlm.nih.gov/pubmed/26595473
Significant interactions with CSA emerged for MGLL at the gene level (p = .009), and for rs604300 within MGLL (ΔR2 = .007, p < .001), the latter of which survived SNP-level Bonferroni correction and was significant in an additional sample with similar directional effects (N = 859; ΔR2 = .005, p = .026). Furthermore, in a third sample (N = 312), there was evidence that rs604300 genotype interacts with early life adversity to predict threat-related basolateral amygdala habituation, a neural phenotype linked to the eCB system and addiction (ΔR2 = .013, p = .047). Rs604300 may be related to epigenetic modulation of MGLL expression. These results are consistent with rodent models implicating 2-arachidonoylglycerol (2-AG), an endogenous cannabinoid metabolized by the enzyme encoded by MGLL, in the etiology of stress adaptation related to cannabis dependence
Carracedo A, Gironella M, Lorente M, Garcia S, Guzman M, Velasco G, Iovanna J. 2006. “Cannabinoids induce apoptosis of pancreatic tumor cells via endoplasmic reticulum stress-related genes” Cancer Res 66: 6748
http://cancerres.aacrjournals.org/content/66/13/6748.long
Pancreatic adenocarcinomas are among the most malignant forms of cancer… the fourth leading cause of cancer death in the United States and the fifth in the Western world overall
cannabinoid receptors are expressed in human pancreatic tumor cell lines and tumor biopsies at much higher levels than in normal pancreatic tissue
cannabinoid administration (a) induced apoptosis, (b) increased ceramide levels, and (c) up-regulated mRNA levels of the stress protein p8. These effects were prevented by blockade of the CB2 cannabinoid receptor or by pharmacologic inhibition of ceramide synthesis de novo
Cannabinoids also reduced the growth of tumor cells in two animal models of pancreatic cancer. In addition, cannabinoid treatment inhibited the spreading of pancreatic tumor cells. Moreover, cannabinoid administration selectively increased apoptosis and TRB3 expression in pancreatic tumor cells but not in normal tissue. In conclusion, results presented here show that cannabinoids lead to apoptosis of pancreatic tumor cells via a CB2 receptor and de novo synthesized ceramide-dependent up-regulation of p8 and the endoplasmic reticulum stress–related genes ATF-4 and TRB3
cannabinoids exert a strong inhibitory effect on the spreading of pancreatic tumor cells not only to adjacent locations such as spleen but also to distal tissues such as liver, diaphragm, stomach, and intestine, thus suggesting that these agents may also decrease the propagation of pancreatic tumor cells.
Although the pancreatic tumor biopsies and cell lines analyzed expressed both CB1 and CB2 cannabinoid receptors, our findings indicate that the CB2 receptor is the one that plays a major role in the proapoptotic effect of cannabinoids in these cells
CB2 receptor is involved in the antitumoral effect of cannabinoids in gliomas ( 9, 19), skin carcinomas ( 11), lymphomas ( 18), and prostate carcinomas ( 30)
p8 and ATF-4 up-regulation mediates cannabinoid-induced apoptosis via induction of the proapoptotic protein TRB3 ( 29, 31)
cannabinoid treatment does not seem to activate this pathway in normal pancreas or spleen, suggesting that these agents may activate the endoplasmic reticulum stress proapoptotic pathway selectively in tumor cells
Carrey N, Butter H, Persinger M, Bialik R. 1995. “Physiological and cognitive correlates of child abuse” J Am Acad Child Adolesc Psychiatry 34(8): 1067-75
http://www.ncbi.nlm.nih.gov/pubmed/7545147/ in dropbox
Verbal and Full Scale IQ scores were inversely related to the severity of abuse that had been experienced. When these variables were used in a discriminant function analysis, children were assigned to the correct group 86% of the time
abuse as delaying cognitive development and inhibiting physiological responsiveness to the environment — 13 full scale IQ gap between abused and reference group
Carrion V, Weems C, Eliez S, Patwardhan A, Brown W, Ray R, Reiss A. 2001. “Attenuation of frontal asymmetry in pediatric posttraumatic stress disorder” Biol Psychiatry 50(12): 943-51
http://www.ncbi.nlm.nih.gov/pubmed/11750890
Casadio P, Fernandes C, Murray R, Di Forti M. 2011. “Cannabis use in young people: The risk for schizophrenia” Neuroscience and Biobehavioral Reviews 35: 1779-1787
http://www.ncbi.nlm.nih.gov/pubmed/21530584 add to dropbox
Until recently, the main types of cannabis available on the “street” were marijuana (grass) and resin (hash) but in recent years a more potent variant termed sinsemilla or skunk has become available in many countries
repeated use of cannabis produces a prolonged and excessive stimulation of the CB1 receptor and disrupts the system
polymorphisms in the AKT1 gene could be involved in cannabis induced psychosis possi-bly through a mechanism of cannabinoid-regulated AKY1/GSK-3 signalling downstream of the dopamine D2 receptor
Other environmental risk factors can also be important. Harley et al. (2010) in studying the effects of childhood trauma, confirmed that cannabis use and childhood trauma were independently asso-ciated with the risk of psychotic symptoms (only cannabis use: OR = 1.9, 95% CI = 0.04–16.5, p = 0.55; only trauma: OR = 2.6, 95% CI = 0.25–14.6, p = 0.23; trauma and cannabis use: OR = 20.9, 95% CI = 2.3–173.5, p = 0.00), but that the joint presence of these two risk factors increased the likelihood of psychotic symptoms in ado-lescence to a much greater extent than would be expected if each risk factor were working independently.
predisposition for psychosis at baseline did not significantly predict cannabis use at follow-up, thus refuting the self-medication hypothesis
Possible explanations for the greater risk in those who start cannabis use early include:
– This association reflects an increased propensity of young peopel with psychotic experiences to commence cannabis use (reverse causality);
– Higher cumulative exposure to cannabis of early users;
– Increased vulnerability to THC during critical phases of brain maturation, such as in early puberty, is reflected in a specific association between psychotic experiences and young inital age of THC exposure
The ability of THC to induce dopamine release in the striatum suggests that THC shares addictive properties with other drugs of abuse, as dopamine has a central role in their rewarding effects, but the increase was modest compare to that obtained with other drugs like cocaine, nicotine or alcohol
While cannabis does not have long term adverse effects for the majority of users, it is clear from studies of psychosis that some individuals are more vulnerable to its effects than others, and that the degree of cannabis exposure and age of first use amplify the harmful effects of cannabis
Caspi A, Moffitt T, Cannon M, McClay J, Murray R, Harrington H, Taylor A, Arseneault L, Williams B, Braithwaite A, Poulton R, Craig I. 2005. “Moderation of the effect of adolescent-onset cannabis use on adult psychosis by a functional polymorphism in the catechol-O-methyltransferase gene: longitudinal evidence of a gene X environment interaction” Biol Psychiatry 57(1): 1117-27
http://www.ncbi.nlm.nih.gov/pubmed/15866551/
functional polymorphism in the catechol-O-methyltransferase (COMT) gene moderated the influence of adolescent cannabis use on developing adult psychosis. Carriers of the COMT valine158 allele were most likely to exhibit psychotic symptoms and to develop schizophreniform disorder if they used cannabis. Cannabis use had no such adverse influence on individuals with two copies of the methionine allele
Casswell, S. 1975. “Cannabis intoxication: effects of monetary incentive on performance, a controlled investigation of behavioural tolerance in moderate users of cannabis” Perceptual and Motor Skills 41: 423-434
http://www.amsciepub.com/doi/pdf/10.2466/pms.1975.41.2.423
Dose-related impairment was found on four of the five tasks, supporting previous findings of cannabis-induced impairment of short-term memory, goal-directed behavior and choice reaction times. Results for three of the tasks suggested that the performance of the motivated subjects was Less affected by the drug than was the performance of the nonmotivated subjects
Both behaviour and experiences while intoxicated have been shown experimentally to be influenced by non-pharmacological factors. Jones demonstrated the importance of the subjects’ expectations and the interpersonal situation in which the drug is taken, for the subjective effects experienced. Experimental manipulation of subjects’ expectancies about the potency of the cannabis they smoked has also resulted in significantly different performance levels following a moderate dose of cannabis (Carlin, Bakker, Halpern, & Post, 1972 )
Users report that they have some control over the degree to which they are involved in the subjective effects and can ‘come down’ and perform normally when necessary (Le Dain Report, 1972). This suggests that the subject’s motivation in experimental investigations of behavioural tolerance to cannabis effects is of crucial importance. It may be unrealistic to assume that incentive to perform the required tasks will be intrinsic to the usual experimental situation, whereas the necessary incentive may be intrinsic to self-set goals during intoxication
Twenty-four of the volunteer subjects were experienced users of cannabis (median frequency of use was three times per wk. and median length of use was 3 yr.), and 24 were non-users all of whom were experienced cigarette smokers and thus able to inhale smoke. Half of each group of 24 was assigned to the motivated group and half to the non-motivated group. The four groups did not differ significantly in mean age, education, and use of alcohol and prescription drugs. Each group contained seven males and five females, five university graduates and seven subjects without university education
cigarettes containing 700 mg of active leaf material, 300 mg of active leaf, or 700 mg of placebo material were smoked. The total leaf content of all cigarettes was 700 rng and in the low dose, placebo and leaf material were thoroughly mixed [MV: leaf? Did they not know that the flowers are the good part?]
it was estimated that the high dose cigarette delivered approximately 4 mg A9-THC and the lower dose delivered approximately 2 mg
Digit Symbol Substitation Task.-A three-way analysis of variance on scores showed a significant (9 < 0.001) dose-related decrement in performance following the administration of cannabis.
Meyer, et al. ( 197 1) allowed ad lib. smoking of cannabis (average dose was 3.6 mg A9-THC) and found mild but non-significant effects on the performance of both casual and heavy users
A third investigation (Miller, Hansteen, Adamec, & Lehmann, 1972) which found no impairment on digit subsritution following a dose and method of administration comparable to the present study did not measure performance until 1% to 2 hr. after smoking
Weil, ed al., while unable to make a definitive comparison, described their naive subjects as ‘grossly impaired’ following the high dose of cannabis (18 mg of ~9-THC was smoked) whereas the chronic users improved slightly after the same dose. Jones ( 1971 ) similarly found no significant change in the performance of frequent users after smoking cannabis containing 9 mg h9-THC
Continuogs Performance Task.-The data obtained from this task were not suitable for parametric analysis. Using the Friedman two-way analysis by ranks (Siegel, 1956), performance by all groups showed no significant effects of drug treatment on either errors of omission or errors of commission… agrees with the results of Weil, et al. ( 1968) who found no cannabis-induced impairment in their naive subjects on this task with or without strobe light distraction. The attention required for adequate performance was not impaired by cannabis
During the early phase of intoxication, in the relatively stimulating circumstances of this experimental administration, and when only short periods of sustained attention are required, socially relevant doses of cannabis seem to induce the “confusion” type of error rather than lapses in attention and are similar in this respect ro the effects of LSD and barbiturates (Mirsky & Kornetsky, 1964). However, lapses in attention may occur more frequently following cannabis consumption during longer lasting vigilance tasks (Moskowitz, Sharma, & Schapero, 1972; Casswell & Marks, 1973a). The relatively long response time allowed in the continuous performance task (1 sec.) will obscure very brief attention lapses induced by cannabis
Goal-directed Serial Alternation nnd Serial Subtraction Tasks A three-way analysis of variance on the transformed serial alternation scores gave a significant (p < 0.05 ) effect of dose on performance, no effect of prior experience with the drug and no statistically significant effect of motivation
None of the differences between the scores of the non-motivated subjects in the placebo condition and the scores of the motivated subjects after smoking cannabis, was statistically significant on either goal-directed serial alternation or serial subtraction
The groups with the best initial baseline performance, i.e., subjects with a university education and males, are those who when motivated are best able to avoid the dose-related impairment found in other groups
greater level of impairment of the motivated subjects in the placebo condition on the most complex of the tasks used in this study, the goal-directed serial alternation. The incentive, in the absence of the counteracting effect of cannabis, has apparently raised the level of arousal beyond the optimum for efficient performance.
This increased inconsistency substantiates the suggestion that cannabis-induced perceptual deficits result from brief lapses of attention (Moskowitz, 1973)
The results of this investigation suggest that when incentive is provided, some cannabis-intoxicated subjects are able to avoid the usual level of impairment in performance of tasks involving short-term memory and goal-directed behaviour. No support was found for the hypothesis that this is true for the moderate users of cannabis to a greater degree than the previously naive subjects
Ceccarini J, De Hert M, Van Winkel R, Peuskens J, Bormans G, Kranaster L, Enning F, Koethe D, Leweke F, Van Laere K. 2013. “Increased ventral striatal CB1 receptor binding is related to negative symptoms in drug-free patients with schizophrenia” Neuroimage 79: 304-12
http://www.ncbi.nlm.nih.gov/pubmed/23624489
Compared to CON, there was a significant increase of CB1 receptor binding in SCZ patients in the nucleus accumbens, insula, cingulate cortex, inferior frontal cortex, parietal and mediotemporal lobe. Furthermore, in the SCZ-F group only, CB1 receptor binding was negatively correlated to negative symptoms and to depression scores, especially in the nucleus accumbens
Centonze D, Battistini L, Maccarrone M. 2008. “The endocannabinoid system in peripheral lymphocytes as a mirror of neuroinflammatory diseases” Curr Pharm Des 14(23): 2370-42
http://www.ncbi.nlm.nih.gov/pubmed/18781987
During immuno-mediated attack of the brain, activation of endocannabinoids represents a protective mechanism, aimed at reducing both neurodegenerative and inflammatory damage through various and partially converging mechanisms that involve neuronal and immune cells
we discuss the appealing working hypothesis that the presence of type-1 cannabinoid receptors on the luminal side, and that of type-2 cannabinoid receptors on the abluminal side of the blood-brain barrier, could drive a unidirectional transport of AEA in the luminal –> abluminal direction (i.e., from blood to brain), thus implying that blood may be a reservoir of AEA for the brain. On this basis, it can be expected that an unbalance of the endogenous tone of AEA in the blood may sustain a similar unbalance of its level within the brain, as demonstrated in Huntington’s disease, Parkinson’s disease, multiple sclerosis, attention-deficit/hyperactivity disorder, schizophrenia, depression and headache
Chait L, Perry J. 1994. “Acute and residual effects of alcohol and marijuana, alone and in combination, on mood and performance” Psychopharmacology 115: 340-349
http://link.springer.com/article/10.1007%2FBF02245075#page-1 add to dropbox
Fourteen male and female subjects were each studied under four conditions: alcohol alone~ marijuana alone, alcohol and marijuana in combination, and no active treatment. Mood and performance assessments were made during acute intoxication and twice the following day (morning and mid-afternoon). Acutely, each drug alone produced moderate levels of subjective intoxication and some degree of behavioral impairment. The drug combination produced the greatest level of impairment on most tasks and “strong” overall subjective ratings. There were few significant interactions between the two drugs, indicating that their effects tended to be additive. Only weak evidence was obtained for subjective or behavioral effects the day after active drug treatments… this study provided little evidence that moderate doses of alcohol and marijuana, consumed either alone or in combination, produce behavioral or subjective impairment the following day
14 subjects (10 males, 4 females) ranged in age from 21 to 34 years (mean = 24.5). All but four reported use of marijuana on at least 100 occasions (lifetime). At the time of participation, subjects reported drinking an average of 7 alcoholic drinks per week (range, 2–12) and smoking marijuana an average of 4 times a month (range, 1-16). Four of the 14 subjects also smoked tobacco cigarettes; all smoked less than a pack a day.
All subjects received four puffs of active marijuana and one alcoholic drink during the practice session
Four treatment conditions were scheduled on the overnight sessions: alcoholic beverage and active marijuana (AM), alcoholic beverage and placebo marijuana (AP), placebo beverage and active marijuana (PM), and placebo beverage and placebo marijuana (PP). Each subject was tested once under each condition and the order of treatments was counterbalanced across subjects… sessions were scheduled 1 week apart on the same day of the week. Subjects were tested in pairs, but treatment condition varied for each member of a pair. Both subjects and experimenter were blind to the treatment conditions
They arrived at 5 P.M. and ate dinner (Fig. 1). At 6:30 P.M. subjects left the CRC and were brought to a room in a separate area of the hospital where behavioral testing and drug administration took place. Subjects were returned to the CRC at 11:45 P.M. Nursing staff ensured that subjects were in their rooms with lights out by midnight, and that subjects were awakened at 7:30 the following morning. Subjects were allowed 30 min to wash and dress before behavioral testing began (about 8 A.M.). Eating, smoking, and coffee drinking were not allowed, however, until after the morning testing was completed. Subjects were tested once more in the afternoon at 3:00 P.M. Subjects were discharged at 4 P.M. Both the morning and afternoon testing were conducted in the same room as the evening testing
Pre-rolled marijuana cigarettes were supplied by the National Institute on Drug Abuse (NIDA). The delta-9-THC contents of the cigarettes were 0.0% (placebo) or 3.6% (active)
95% ethanol in diet tonic water and lime juice. The content (v/v) of the ptacebo beverage was 1% ethanol (as a taste mask), 4% lime juice, and 95% tonic, while the content of the alcoholic beverage was 10% ethanol, 4% lime juice, and 86% tonic. Drink volume was adjusted for gender and body weight so that the dose for the alcohol conditions was 0.6 g/kg for males and 0.5 g/kg for females. Drink volume was 540 ml/70 kg for males and 450 ml/70 kg for females. The beverages were served cold in 1 1 plastic containers
The effect of active marijuana on heart rate varied considerably across subjects, from no increase at all to a 56 bpm increase. Neither placebo marijuana nor alcohol significantly altered heart rate, and there was no interaction between marijuana and alcohol
The only significant effect for ratings of drug liking was a main effect of Marijuana [F(1, 13) = 9.9, P < 0.01]. Subjects rated liking active marijuana (mean = 64.4) more than placebo (mean = 51.3
The POMS was insensitive to the drug treatments, Mthough five of the eight scales did show significant main effects of Time, reflecting more negative mood and greater fatigue as the evening progressed, regardless of drug condition
Four of the VAS scales were sensitive to the drug treatments (Fig. 2). Ratings of “high” were increased only by marijuana [Marijuana x Time: F(4; 52) = 17.5, P < 0.0001] whereas ratings of “drunk” were increased only by alcohol [Alcohol x Time: F(4, 32)= 8.7, P = 0.0005]. With active marijuana, “high” ratings peaked after the first drug administration and showed little further increase after the second drug administration. In contrast, ratings of “drunk” continued to increase throughout the evening when alcohol was given. Ratings of”stimutated” were increased by both drugs [Alcohol x Time: F(4, 52) = 3.4, P < 0.025; Marijuana x Time: F(4, 52) = 3.4, P < 0.05]. Marijuana, but not alcohol, also resulted in higher ratings of”hungry” [Marijuana x Time: F(4, 52) = 6.4, P < 0.005]. Interestingly, the effects of marijuana on “hungry” ratings showed a different time course from the effects of the drug on “high,” with “hungry” ratings peaking later in the session (Fig. 2)
Both drugs affected time production [Alcohol x Target Interval: F(2, 24) = 9.7, P < 0.01; Marijuana x Target Interval: F(2, 24) = 4.2, P < 0.05]. As shown in Fig. 3, the two drugs produced opposite effects – alcohol administration resulted in subjects producing time intervals longer than the targets (overproduction), whereas marijuana administration occasioned underproduction. When the two drugs were given in combination, (AM condition) the effects canceled each other out, resulting in no effect.
Standing steadiness. Only alcohol impaired performance on this task [Alcohol x Time: F(1, 13) = 9.9, P < 0.01]. The mean score (time summed for both legs) dropped from 57.7 s before drug to 45.9 s after drug.
DSST. Alcohol impaired performance on all measures of DSST performance: number attempted [Alcohol x Time: F(1, 13) = 23.3, P < 0.0005], number correct [Alcohol x Time: F(1, 13) = 22.3, P < 0.0005], and percentage correct [Alcohol x Time: F(1, 13) = 7.2, P < 0.025]. Number correct decreased from a mean of 43.9 before alcohol to 37.4 after, while mean percentage correct decreased from 96.5 to 91.1. Marijuana affected only percentage correct [Marijuana x Time: /7(1, 13) = 8.3, P < 0.025], which declined from a mean of 96.t to 91.3.
Backward digit span. Only alcohol affected this task [Alcohol x Time: F(1, 13) = 29.0, P < 0.0001]. The average span was 6.5 digits before alcohol, compared with 5.1 digits after alcohol.
Logical reasoning. No significant drug effects were obtained on this task.
Divided attention. There were no effects of the drug treatments on either hit rate or mean reaction time for responding to the primary target. Alcohol increased errors in estimating the number of occurrences of the secondary target [Alcohol x Time: F(1, 13) = 7.3, P < 0.025], from a mean of 1.0 errors before drug to 1.6 errors after drug. Marijuana increased false alarm responses (responding to the secondary target as if it were the primary target) from 0.9 to 1.6, before and after drug, respectively [Marijuana x Time: F(1, 13)= 5.9, P < 0.05].
Free recall. Only alcohol affected this task, producing a decrement in the number of words recalled from a mean of 9.3 before drug to 6.6 after drug [Alcohol x Time: F(t, 13) = 23.5, P < 0.0005]
Sleep questionnaire. Three of the four factors measured by the LSEQ showed significant drug-related effects (Table 1). Subjects rated it easier to get to sleep (GTS) after both drugs [Alcohol: F(1, 13) — 15.8, P < 0.0025; Marijuana: F(1, 13) = 6.4, P < 0.025]. For ratings of quality of sleep (QOS), a significant Alcohol x Marijuana interaction was obtained [F(1, 13) — 4.9, P < 0.05]. As Table 1 indicates, this interaction was due to higher ratings of sleep quality when alcohol and marijuana were administered alone, but relatively lower ratings after the drug combination. Finally, behavior following wakefulness (BFW) scores were decreased by Alcohol [F(1, 12) = 7.4, P < 0.025], indicating subjects felt more tired and clumsy
The present study yielded little evidence for next-day effects after any of the active drug treatments.
none of the apparent after-effects obtained in our previous studies was replicated here… In our first study (Chait et al. 1985) we found increased scores on the A and BG scales of the ARCI, and overproduction on the time production task, the morning after active marijuana. In the second study (Chait 1990), we observed underproduction on the time production task, impaired performance on backward digit span, and slowing of reaction time on the divided attention task the morning after active marijuana… type I errors.
One effect of marijuana that did replicate here, although not strictly speaking a next-day effect, was its effect on subjective sleep: in both the present and the previous study in which the LSEQ was used (Chait 1990) active marijuana increased subject ratings of ease of getting to sleep (Table 1); the mean increases in the two studies were of comparable magnitude (12-15 units).
ur failure to find significant behavioral impairment the day after this dose of alcohol is not surprising, since most studies that have reported impairment the day after drinking used doses higher than 1.2 g/ kg (Seppala et al. 1976; Myrsten et al. 1980), and some studies using higher doses found no reliable evidence of impairment (Takala et al. 1958)
During the evening sessions, heart rate increased more after marijuana smoking during the first compared with the second smoking period. Similarly, ratings of “high” increased sharply after the first, but not the second smoking period (Fig. 2). Since CO boost did not differ between the two smoking periods, these results provide strong evidence for acute tolerance to these effects of marijuana, a phenomenon that has been observed by others (Coc- chetto et al. 1981; Perez-Reyes et al. 1981).
inability to show marijuana-induced increases in hunger in at least some of these prior studies (Chait et al. 1985, 1988a,b; Zacny and Chait 1989, t991; Zacny and de Wit 1989, 1991; Chait and Zacny 1992) may have been because the effect is delayed, not reaching a peak until 1 h after smoking.
moderate doses of these two commonly used substances have little effect on mood or performance the day after use, and that at least equal attention should be paid to time of day as an important influence upon human behavior
Chakraborty A, Anstice N, Jacobs R, LaGasse L, Lester B, Wouldes T, Thompson B. 2015. “Prenatal exposure to recreational drugs affects global motion perception in preschool children” Nature: Scientific Reports 5: 16921
http://www.nature.com/articles/srep16921
Global motion perception was measured in one hundred and forty-five 4.5-year-old children who had been exposed to different combinations of methamphetamine, alcohol, nicotine and marijuana prior to birth and 25 unexposed children. Self-reported drug use by the mothers was verified by meconium analysis. We found that global motion perception was impaired by prenatal exposure to alcohol and improved significantly by exposure to marijuana. Exposure to both drugs prenatally had no effect
Demographically the cohort was 52.5% European, 36.5% Māori and 11% other. The children had been exposed to a range of different drugs: 75.2% to nicotine, 56.4% to alcohol, 44.2% to methamphetamine and 40% to marijuana. The majority of children (81.3%) had been exposed to multiple drugs. Twenty-five children (15%) had no drug exposure and provided a non-drug exposed comparison group
Only alcohol and marijuana exposure had independent effects on global motion perception after controlling for the effects of multiple drug exposure, verbal IQ, ethnicity, habitual visual acuity, stereoacuity, and sex. Children who were exposed prenatally to alcohol had elevated (worse) motion coherence thresholds (Fig. 1A) whereby global motion perception was significantly poorer (t163 = −2.26, p = 0.002) than that of children not exposed to alcohol (alcohol exposed, n = 95: 59 ± 21% motion coherence threshold vs alcohol non-exposed, n = 70: 50 ± 23%). Unexpectedly, children exposed to marijuana had significantly lower (better) motion coherence thresholds (Fig. 1B) (t163 = 3.52, p = 0.001) than those of children not exposed to marijuana [marijuana exposed, n = 67: 46 ± 20% vs marijuana non-exposed, n = 98: 63 ± 25%]. A significant interaction between the effects of alcohol exposure and marijuana exposure on motion coherence thresholds was also present (Fig. 1C, F1,114 = 7.7, p = 0.006), whereby exposure to marijuana in the absence of alcohol was associated with improved global motion perception (mean motion coherence threshold = 34 ± 11%, n = 20), which was significantly lower (better) (t41 = 4.42, p < 0.001) than that of children who had not experienced prenatal drug exposure (mean motion coherence threshold = 58 ± 23%, n = 25). However global motion perception for children exposed to both marijuana and alcohol (mean motion coherence threshold = 53 ± 24%, n = 48) was no different (t70 = −0.28, p = 0.39) from that of children who had no drug exposure (mean motion coherence threshold = 58 ± 23%, n = 25)
visual acuity (measured with habitual refractive correction if worn), stereoacuity, and verbal IQ (measured with Wechsler Preschool and Primary Scale of Intelligence-III) were unaffected by drug exposure in this group of children
More frequent maternal use of marijuana (Fig. 2a) during pregnancy and more joints smoked per occasion (Fig. 2b) were associated with lower motion coherence thresholds, indicating better global motion perception. The opposite of this was true for children (n = 46) exposed to alcohol
none of the children in this study had been diagnosed with foetal alcohol syndrome. This suggests that prenatal exposure to levels of alcohol that are not sufficient to induce foetal alcohol syndrome can still impair the cortical processing of visual information as assessed using a behavioural measure of global motion perception
interesting possibility that prenatal marijuana exposure may improve dorsal stream function but impair ventral stream function. Alternatively, the detrimental effects of marijuana on visual processing reported in previous studies may reflect impairments at the level of visual attention, visual memory, or response inhibition rather than visual perception [MV: or may be artifacts of bad study designs and confounding by other factors]
Marijuana neutralizes the effect of nicotine in an animal model of addiction54
Chang L, Yakupov R, Cloak C, Ernst T. 2006. “Marijuana use is associated with a reorganised visual-attention network and cerebellar hypoactivation” Brain 129(5): 1096-1112
http://brain.oxfordjournals.org/content/129/5/1096.long
24 chronic MJ users, 12 abstinent, 12 active; 19 age-, sex- and education-matched controls age of first marijuana use was 14.7 ± 0.4 (12–16) years for the THC(−) subjects and 15.5 ± 0.9 (9–20) years for the THC(+)
decreased activation in the right prefrontal, medial and dorsal parietal, and medial cerebellar regions, but greater activation in various frontal, parietal and occipital brain regions during the visual-attention tasks
Active marijuana users, with positive urine tests for THC, showed greater activation in the frontal and medial cerebellar regions than abstinent
earlier age of first use and greater estimated cumulative dose of THC exposure were related to lower BOLD signals in the right prefrontal region and medial cerebellum
| CES-depression scale |
6.90 ± 0.98 |
9.42 ± 1.92 |
12.67 ± 2.71 |
0.08* |
“minimal depressive symptoms”
smoke 27.9 days per month, 6047 ± 2908 joints
THC(+) marijuana user group showed better performance on the response reversal/visual scanning tests and a trend for faster reaction times on the choice reaction task
Marijuana users performed better while tracking 2 balls, but not when tracking 3 or 4
Chavarría-Siles I, Contreras-Rojas J, Hare E, Walss-Bass C, Quezada P, Dassori A, Contreras S, Medina R, Ramírez M, Salazar R, Raventos H, Escamilla MA. 2008. “Cannabinoid receptor 1 gene (CNR1) and susceptibility to a quantitative phenotype for hebephrenic schizophrenia” Am J Med Genet B Neuropsychiatr Genet 147(3):279-84
http://www.ncbi.nlm.nih.gov/pubmed/18186055
subjects with schizophrenia who had an elevated lifetime dimensional score for negative and disorganized symptoms, which we used as a proxy for “hebephrenia.”
we found association of these hebephrenic subjects and the (AAT)n-repeat marker of the CNR1 (multi-allelic P = 0.0368). Our hypothesis that an association with the (AAT)n-repeat marker of CNR1 would not be found with the more general type of schizophrenia was also confirmed. Schizophrenic subjects with prominent lifetime scores for disorganization and negative symptoms (dimension for hebephrenia) are associated with the CNR1 gene and present a type of symptomatology that resembles chronic cannabinoid-induced psychosis
Cheetham A, Allen N, Whittle S, Simmons J, Yucel M, Lubman D. 2012. “Orbitofrontal volumes in early adolescence predict initiation of cannabis use: a 4-year longitudinal and prospective study” Biol Psychiatry 71(8): 684-92
http://www.ncbi.nlm.nih.gov/pubmed/22129756
Although these changes are frequently attributed to the neurotoxic effects of cannabis, it is possible that some abnormalities might predate use and represent markers of vulnerability
Smaller orbitofrontal cortex volumes at age 12 years predicted initiation of cannabis use by age 16 years. The volumes of other regions (amygdala, hippocampus, and anterior cingulate cortex) did not predict later cannabis use
Chen X, Williamson V, An S-S, Hettema J, Aggen S, Neale M, Kendler K. 2008. “Cannabinoid Receptor 1 Gene Association with Nicotine Dependence” Arch Gen Psychiatry 65(7): 816-824
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2733353/
Chhatwal J, Davis M, Maguschak K, Ressler K. 2005. “Enhancing cannabinoid neurotransmission augments the extinction of conditioned fear” Neuropsychopharmacology 30(3):516-24
http://www.ncbi.nlm.nih.gov/pubmed/15637635
CB1 antagonist rimonabant (SR141716A) prior to extinction training led to significant, dose-dependent decreases in extinction
CB1 agonist WIN 55,212-2 did not appear to affect extinction, administration of AM404, an inhibitor of eCB breakdown and reuptake, led to dose-dependent enhancements in extinction
AM404-treated animals showed decreased shock-induced reinstatement of fear; could not be attributed to alterations in the expression of conditioned fear, locomotion, shock reactivity, or baseline startle, as these parameters seemed unchanged by AM404
coadministration of rimonabant with AM404 blocked this enhancement
eCB modulators may be therapeutically useful as adjuncts for exposure-based psychotherapies such as those used to treat Post-Traumatic Stress Disorder and other anxiety disorders
de Chiara V, Errico F, Musella A, Rossi S, Mataluni G, Sacchetti L, Siracusano A, Castelli M, Cavasinni F, Bernardi G, Usiello A, Centonze D. 2010. “Voluntary exercise and sucrose consumption enhance cannabinoid CB1 receptor sensitivity in the striatum” Neuropsychopharmacology 35(2): 374-387
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3055381/
Running wheel and sucrose consumption have rewarding and reinforcing properties in rodents, and share many neurochemical and behavioral characteristics with drug addiction… . We found that cannabinoid CB1 receptor-mediated presynaptic control of striatal inhibitory postsynaptic currents was remarkably potentiated after these environmental manipulations… the sensitivity of glutamate synapses to CB1 receptor stimulation was unaltered, as well as that of GABA synapses to the stimulation of presynaptic GABAB receptors
the upregulation of cannabinoid transmission induced by wheel running or sucrose had a crucial role in the protective effects of these environmental manipulations against the motor and synaptic consequences of stress
Chiu Y-C, Egner T. 2015. “Inhibition-induced forgetting: when more control leads to less memory” Psychological Science 26(1): 27-38
http://www.ncbi.nlm.nih.gov/pubmed/25398560
hypothesis that inhibition-induced forgetting occurs when response inhibition shunts attentional resources from perceptual stimulus encoding to action control.
Chiurchiù V, Leuti A, Maccarrone M. 2015. “Cannabinoid signaling and neuroinflammatory diseases: a melting pot for the regulation of brain immune responses” Journal of Neuroimmune Pharmacology 10(2): 268-280
http://link.springer.com/article/10.1007/s11481-015-9584-2#/page-2
Cannabis sativa-derived phytocannabinoids, as well as synthetic cannabinoids, are endowed with significant immunoregulatory and anti-inflammatory properties, both in peripheral tissues and in the CNS, through the activation of cannabinoid receptors. In this review, the immunomodulatory effects of cannabinoid signaling on the most relevant brain immune cells will be discussed. In addition, the impact of cannabinoid regulation on the overall integration of the manifold brain immune responses will also be highlighted, along with the implication of these compounds as potential agents for the management of neuroinflammatory disorders
Choi J, Jeong B, Rohan M, Polcari A, Teicher M. 2009. “Preliminary evidence for white matter tract abnormalities in young adults exposed to parental verbal abuse.” Biol Psychiatry 65(3):227-34
http://www.ncbi.nlm.nih.gov/pmc/articles/pmid/18692174/
Three WM tract regions had significantly reduced FA: 1) arcuate fasciculus in left superior temporal gyrus, 2) cingulum bundle by the posterior tail of the left hippocampus, and 3) the left body of the fornix.
Across groups, FA in region 1 correlated with verbal IQ and verbal comprehension index. Fractional anisotropy in region 2 was inversely associated with ratings of depression, dissociation, and limbic irritability. Fractional anisotropy in region 3 was inversely correlated with ratings of somatization and anxiety.
Chuchawankul S, Shima M, Buckley N, Hartmann C, McCoy K. 2004. “Role of cannabinoid receptors in inhibiting macrophage costimulatory activity” Int Immunopharmacol 4(2): 265-78
http://www.ncbi.nlm.nih.gov/pubmed/14996418/
THC decreased costimulation provided by peritoneal cells elicited with polystyrene beads and thioglycollate, but the drug had no influence with macrophages elicited with thioglycollate alone. Bead administration induced CB2 mRNA expression in macrophages, while CB1 mRNA was not detected. Although inhibition was associated with functional heat-stable antigen, a costimulatory molecule, on macrophages, THC exposure did not alter cell surface heat-stable antigen expression. Inhibition by THC and anti-heat-stable antigen antibody was not additive suggesting the inhibitory mechanisms may overlap… CB1-selective antagonist SR141716A completely reversed, and CB2-selective antagonist SR144528 partially blocked THC’s inhibition
High affinity synthetic agonist CP55,940, but not partial agonist THC, impaired costimulation by macrophages from mice lacking CB2 receptor
Churchwell J, Lopez-Larson M, Yurgelun-Todd D. 2010. “Altered frontal cortical volume and decision making in adolescent cannabis users” Front Psychol 1: 225
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3153830/
fails to control for trauma which separately found same effect; small sample size, and two outliers produce the correlation between low MoPFC and age of first use; Axis I diagnoses excluded but CA had subclinical levels of pathology and were not asked about why they were using
no significant correlations between right moPFC volume and several measures of cannabis use, total moPFC volume was positively associated with age of first use
18 cannabis abusing adolescents (CA; 2 female and 16 male subjects; mean age, 17.7 years; range 16–19 years), and 18 healthy controls (HC; 6 female and 12 male subjects; mean age, 17.2 years; range 16–19 years)
cannabis abusing adolescents have decreased right moPFC volume compared to controls, p =0.01, d=0.92, CI0.95=0.21, 1.59
=0.01, d=0.92, CI0.95=0.21, 1.59
total moPFC volume was positively correlated with age of first use r (18)=0.49, p<0.03
Morphogenesis of the prefrontal cortex has been associated with an enhanced capacity to temporally organize action plans and choices as a means of reaching goals
decreased right moPFC cortical volume was also shown to be associated with increased observed impulsivity in a sample of adolescent males (Boes et al., 2009)
exclusionary criteria: (1) presence of any current Axis I disorder other than cannabis abuse (for the cannabis group); (2) presence of a serious medical illness, including diabetes; (3) presence of a neurological disorder; (4) history of head trauma resulting in loss of consciousness requiring hospital evaluation; (5) history of ECT treatment; (6) estimated IQ<85 (based on measures of verbal fluency and academic history); (7) claustrophobia or metal implanted within the body
Cannabis abusing subjects in the present study performed somewhat better on the verbal fluency test (M=45.8, SD=9.5) than healthy controls (M=37.5, SD=8.7)
no correlation between tetrahydrocannabinol metabolite values and BIS scores, suggesting that active levels did not influence BIS responding
Hamilton Anxiety Rating Scale (HAM-A; HC, M=1.27, SD=1.93; CA, M=2.0, SD=2.66) or depression as assessed with the Hamilton Depression Rating Scale (HAM-D; HC, M=0.77, SD=1.43; CA, M=2.11, SD=3.07)
Cichewicz D, Haller V, Welch S. 2001. “Changes in opioid and cannabinoid receptor protein following short-term combination treatment with delta(9)-tetrahydrocannabinol and morphine” J Pharmacol Exp Ther 297(1): 121-7
http://www.ncbi.nlm.nih.gov/pubmed/11259535
upregulation of CB1 in spinal cord. Coadministration of THC and morphine blocks development of morphine tolerance without reducing antinociceptive effects
Cloitre M, Stolbach B, Herman J, van der Kolk B, Pynoos R, Wang J, Petkova E. 2009. “A developmental approach to complex PTSD: childhood and adult cumulative trauma as predictors of symptom complexity” J Trauma Stress 22(5): 399-408
http://www.pubfacts.com/detail/19795402/A-developmental-approach-to-complex-PTSD:-childhood-and-adult-cumulative-trauma-as-predictors-of-sym
Childhood cumulative trauma but not adulthood trauma predicted increasing symptom complexity in adults. Cumulative trauma predicted increasing symptom complexity in the child sample. Results suggest that Complex PTSD symptoms occur in both adult and child samples in a principled, rule-governed way and that childhood experiences significantly influenced adult symptoms
Cloitre M, Garvert D, Weiss B, Carlson E, Bryant R. 2014. “Distinguishing PTSD, Complex PTSD, and Borderline Personality Disorder: A latent class analysis” Eur J Psychotraumatol. 5
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4165723/
spectrum of diagnoses moving from PTSD to CPTSD and BPD
the following symptoms were highly indicative of placement in the BPD rather than the CPTSD class: (1) frantic efforts to avoid real or imagined abandonment, (2) unstable and intense interpersonal relationships characterized by alternating between extremes of idealization and devaluation, (3) markedly and persistently unstable self-image or sense of self, and (4) impulsiveness. Given the gravity of suicidal and self-injurious behaviors, it is important to note that there were also marked differences in the presence of suicidal and self-injurious behaviors with approximately 50% of individuals in the BPD class reporting this symptom but much fewer and an equivalent number doing so in the CPSD and PTSD classes (14.3 and 16.7%, respectively)
The focus of treatment for BPD concerns reduction of life-interfering behaviors such as suicidality and self-injurious behaviors, a reduction in dependency on others and an increase in an internalized and stable sense of self (e.g., Dialectical Behavior Therapy, Linehan, 1993). In contrast, treatment programs for CPTSD focus on reduction of social and interpersonal avoidance, development of a more positive self-concept and relatively rapid engagement in the review and meaning of traumatic memories (e.g., Cloitre et al., 2006)
Cluny N, Keenan C, Reimer R, Le Foll B, Sharkey K. 2015. “Prevention of diet-induced obesity effects on body weight and gut microbiota in mice treated chronically with Δ9-tetrahydrocannabinol” PLoS One DOI: 10.1371/journal.pone.0144270
http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0144270
THC reduced weight gain, fat mass gain and energy intake in DIO but not lean mice. DIO-induced changes in select gut microbiota were prevented in mice chronically administered THC. THC had no effect on locomotor activity or whole gut transit in either lean or DIO mice
Chronic THC treatment reduced energy intake and prevented high fat diet-induced increases in body weight and adiposity; effects that were unlikely to be a result of sedation or altered gastrointestinal transit. Changes in gut microbiota potentially contribute to chronic THC-induced actions on body weight in obesity
Coccaro E, McCloskey M, Fitzgerald D, Phan K. 2007. “Amygdala and orbitofrontal reactivity to social threat in individuals with impulsive aggression” Biological Psychiatry 62(2): 168-178
http://www.biologicalpsychiatryjournal.com/article/S0006-3223(06)01064-X/abstract
Relative to controls, individuals with IED exhibited exaggerated amygdala reactivity and diminished OFC activation to faces expressing anger. Extent of amygdala and OFC activation to angry faces were differentially related to prior aggressive behavior across subjects. Unlike controls, aggressive subjects failed to demonstrate amygdala-OFC coupling during responses to angry faces
These findings provide evidence of amygdala-OFC dysfunction in response to an ecologically-valid social threat signal (processing angry faces) in individuals with a history of impulsive aggressive behavior
Cohen R, Grieve S, Hoth K, Paul R, Sweet L, Tate D, Gunstad J, Stroud L, McCaffery J, Hitsman B, Niaura R, Clark C, McFarlane A, Bryant R, Gordon E, Williams L. 2006. “Early life stress and morphometry of the adult anterior cingulate cortex and caudate nuclei” Biol Psychiatry 59(10): 975-82
http://www.ncbi.nlm.nih.gov/pubmed/16616722/
Participants with greater than two ACEs had smaller ACC and caudate nuclei than those without ACEs
Cole S, Hawkley L, Arevalo J, Sung C, Rose R, Cacioppo J. 2007. “Social regulation of gene expression in human leukocytes” Genome Biol 8(9): R189
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2375027/
The present study analyzed genome-wide transcriptional activity in people who chronically experienced high versus low levels of subjective social isolation (loneliness) to assess alterations in the activity of transcription control pathways that might contribute to increased adverse health outcomes in social isolates
Promoter-based bioinformatic analyses showed under-expression of genes bearing anti-inflammatory glucocorticoid response elements (GREs; p = 0.032) and over-expression of genes bearing response elements for pro-inflammatory NF-κB/Rel transcription factors (p = 0.011). This reciprocal shift in pro- and anti-inflammatory signaling was not attributable to differences in circulating cortisol levels, or to other demographic, psychological, or medical characteristics. Additional transcription control pathways showing differential activity in bioinformatic analyses included the CREB/ATF, JAK/STAT, IRF1, C/EBP, Oct, and GATA pathways
impaired transcription of glucocorticoid response genes and increased activity of pro-inflammatory transcription control pathways provide a functional genomic explanation for elevated risk of inflammatory disease in individuals who experience chronically high levels of subjective social isolatio
Cole J, Toga A, Hojatkashani C, Thompson P, Costafreda S, Cleare A, Williams S, Bullmore E, Scott J, Mitterschiffthaler M, Walsh N, Donaldson C, Mirza M, Marquand A, Nosarti C, McGuffin P, Fu C. 2010. “Subregional hippocampal deformations in major depressive disorder.” J Affect Disord 126(1-2): 272-7
http://www.ncbi.nlm.nih.gov/pubmed/20392498/
Both right (P=0.001) and left (P=0.005) hippocampal volumes were reduced in patients relative to healthy controls (n=37 patients and n=37 controls), while only the right hippocampus (P=0.016) showed a reduced volume in a subgroup of first-episode depression patients
Compton M, Furman A, Kaslow N. 2004. “Preliminary evidence of an association between childhood abuse and cannabis dependence among African American first-episode schizophrenia-spectrum disorder patients” Drug Alcohol Depend 76(3):311-6
http://www.ncbi.nlm.nih.gov/pubmed/15561482
Schizophrenics with comorbid cannabis dependence reported greater childhood physical and sexual abuse than schizophrenics without cannabis dependence
Compton M, Furman A, Kaslow N. 2004. “Lower negative symptom scores among cannabis-dependent patients with schizophrenia-spectrum disorders: preliminary evidence from an African American first-episode sample” Schizophr Res 71(1): 61-4
http://www.ncbi.nlm.nih.gov/pubmed/15374573
those with cannabis dependence had significantly lower Positive and Negative Syndrome Scale negative subscale scores compared to those without cannabis dependence. The two groups did not differ on postive and general psychopathology subscale scores
Compton M, Kelley M, Ramsay C, Pringle M, Goulding S, Esterberg M, Stewart T, Walker E. 2009. “Association of pre-onset cannabis, alcohol and tobacco use with age at onset of prodrome and age at onset of psychosis in first-episode patients” American Journal of Psychiatry 166(11): 1251-1257
http://psychiatryonline.org/doi/full/10.1176/appi.ajp.2009.09030311
Whereas classifying participants according to maximum frequency of use prior to onset (none, ever, weekly, or daily) revealed no significant effects of cannabis or tobacco use on risk of onset, analysis of change in frequency of use prior to onset indicated that progression to daily cannabis and tobacco use was associated with an increased risk of onset of psychotic symptoms
progression to daily use resulted in a much larger increased relative risk of onset of psychosis in females than in males
12-month prevalence estimates of use and misuse of cannabis were 29% and 19%, respectively, and lifetime use and misuse estimates were 42% and 23%, respectively
In patients with first-episode psychosis, reported rates of cannabis misuse have ranged from 15% to 65% (3 – 11) , rates of alcohol misuse from 27% to 43% (9 , 11 – 15) , and rates of daily cigarette smoking from 50% to 75% (11 , 13 ,16)
one critique of the literature is that the possible influence of cannabis use on prodromal symptoms has not been explored
family history in first-degree relatives was present in only 13.4% of participants, making it difficult to estimate those effects with any reasonable certainty
large percentage of daily cannabis users (40.6%) and daily tobacco users (44.1%) but relatively few daily alcohol users (7.9%)
no significant effects of cannabis or tobacco use on risk of onset of psychotic symptoms but showed an effect of weekly and daily alcohol use on age at onset of psychosis
progression to daily use was associated with a larger increased relative risk for onset of psychosis in female patients (hazard ratio=5.154) than in male patients (hazard ratio=3.359) male patients with no history of use and those with ever or weekly use had higher hazard ratios than the female comparison groups (2.250 compared with 1.000, and 2.329 compared with 2.049, respectively), which was expected, although the male and female patients were not significantly different at those levels. However, for daily users, the hazard ratio was 3.359, compared with 5.154, indicating a reversal of the pattern of findings (higher in female patients)
increased use of cannabis by those imminently approaching the onset of psychotic symptoms is also a reasonable interpretation of these data
Conklin C, Westen D. 2005. “Borderline Personality Disorder in Clinical Practice” Americal Journal of Psychiatry 162(5): 867-875
http://ajp.psychiatryonline.org/doi/full/10.1176/appi.ajp.162.5.867
A random national sample of 117 experienced psychiatrists and psychologists from the membership registers of the American Psychiatric Association and American Psychological Association provided data on a randomly selected patient with borderline personality disorder (N=90) or dysthymic disorder (N=27) from their practice
Gunderson’s review (20)indicated that the axis I disorders most frequently found in borderline personality disorder patient samples are dysthymic disorder, major depression, substance abuse, posttraumatic stress disorder, and eating disorders and that at least one-half of borderline personality disorder patients have major depressive disorder, dysthymia, or both. Although borderline personality disorder has been found to have high rates of comorbidity with virtually all axis II disorders, the highest diagnostic overlap appears to be with histrionic and avoidant personality disorders (20, 21). With regard to adaptive functioning, research findings have associated borderline personality disorder with self-injurious behavior such as skin cutting and burning and with psychiatric hospitalizations, suicidality, difficulty maintaining relationships, and difficulty maintaining appropriate employment
intense dysphoric affect is a core, rather than co-occurring, feature of borderline personality disorder… chronic major depression and chronic feelings of helplessness, hopelessness, worthlessness, guilt, loneliness, and emptiness appear to be central to the disorder
we asked clinicians to select a female patient (to avoid the confounding factor of gender and to maximize power, because 75%–80% of patients who receive a diagnosis of borderline personality disorder are female [20, DSM-IV]
Of the 117 clinician-participants, 19% (N=22) were psychiatrists and 81% (N=95) were psychologists (the latter responded at a much higher rate to the initial solicitation); 42% (N=49) were female… 21% (N=24) describing their psychotherapeutic orientation as cognitive behavioral or behavioral, 44% (N=52) as psychodynamic or psychoanalytic, 32% (N=37) as eclectic, and 3% (N=4) as other.
The borderline personality disorder group distinguished itself both by the sheer number of comorbid diagnoses on average and by the specific diagnoses that have commonly been reported in reviews of studies of borderline personality disorder patients (20, 21).
Most (70%, N=63) of the borderline personality disorder patients had attempted suicide. Attempters on average had made 3.89 attempts (SD=6.72), with the severity of the most dangerous attempt rated on average as “moderate, requiring medical attention.” Most (63%, N=57) of the borderline personality disorder patients had at least one psychiatric hospital admission, and those who had a history of hospitalization had an average of 3.67 admissions (SD=3.92). More than one-half of the borderline personality disorder patients (52%, N=47) had self-injured. Of the 47 borderline personality disorder patients who self-injured, 81% (N=38) cut, 23% (N=11) burned, and 13% (N=6) severely scratched or tore their skin; an additional 26% (N=12) had repeated accidents
41% (N=37) had been in abusive relationships in adulthood, with the majority in the role of victim (60%, N=22), a substantial minority in the roles of both victim and perpetrator (38%, N=14), and only one exclusively in the role of perpetrator. Nearly one-third (32%, N=29) had been the victim of rape or sexual assault in adulthood (see reference 46), and, for borderline personality disorder patients who reported any such incident, rape or sexual assault occurred on average 2.75 times (SD=3.66)
Perhaps most striking, the two items most descriptive of borderline personality disorder patients in both samples were “Tends to feel unhappy, depressed, or despondent” and “Emotions tend to spiral out of control.”
Borderline personality disorder patients appeared to have frequent or chronic minor illnesses, leading to missed appointments, days off from work, visits to the doctor, or subjective distress.
Among the DSM-IV criteria that did not receive high average rankings were items describing the tendency to see people as “all good” or “all bad” (either because empirically these concepts are not as central to the diagnosis or because splitting them into two items may have decreased the ranking of both) and items describing specific forms of impulsivity, such as alcohol abuse and promiscuous sex. The hybrid criterion added to DSM-IV regarding transient psychotic symptoms and dissociative episodes did not rank highly. However, a related item that ranked tenth in the borderline personality disorder sample better appears to capture an aspect of the construct originally intended by Gunderson (21) and should be considered as a replacement criterion for DSM-V: “tends to become irrational when strong emotions are stirred up; may show a noticeable decline from customary level of functioning.”
the data provided by psychologists and by psychiatrists did not show any pattern of differences in this or any of our prior studies using this method, despite substantially different response rates
male and non-Caucasian borderline personality disorder patients are understudied groups, and broader sampling, including oversampling to maximize representativeness of the population, would strengthen future investigations.
Conrad C. 2008. “Chronic stress-induced hippocampal vulnerability: the glucocorticoid vulnerability hypothesis” Rev Neurosci 19(6): 395-411
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2746750/
The hippocampus, a limbic structure important in learniJng and memory, is particularly sensitive to chronic stress and to glucocorticoids
a chronic stress history, which includes repeated elevation of glucocorticoids, may make the hippocampus vulnerable to potential injury
glucocorticoids can compromise the hippocampus by producing dendritic retraction, a reversible form of plasticity that includes dendritic restructuring without irreversible cell death. Conditions that produce dendritic retraction are hypothesized to make the hippocampus vulnerable to neurotoxic or metabolic challenges
A racing heart and high blood pressure triggered by epinephrine to maximize blood flow is critical for assisting in escaping the stressor, but a persistent racing heart and high blood pressure increases the risk of heart failure and arteriosclerosis. Elevated glucocorticoids are essential for the redistribution of energy resources, but the long-term elevation of glucocorticoids starves some tissues of necessary resources and hinders immune function, increasing the susceptibility to disease
Guinea pigs injected daily with glucocorticoids for four weeks exhibited brain damage in the hypothalamus and hippocampus /4
the hippocampus provides negative feedback to the HPA axis, with hippocampal stimulation and lesions causing decreases and increases in HPA axis activity, respectively /36,40,43,58,85
glucocorticoids secreted during periods of stress desensitize the hippocampus to further glucocorticoid exposure by downregulating glucocorticoid receptors, an effect that is self-correcting and therefore reversible. At some point, however, the downregulation of glucocorticoid receptors precipitates further hypersecretion of glucocorticoids until permanent hippocampal cell loss occurs. This irreversible hippocampal damage was proposed to make the hippocampus irreversibly insensitive to further glucorticoid elevations, creating a feed-forward cycle of elevated glucocorticoids and continued hippocampal destruction as an individual ages. While elegant, the glucocorticoid cascade hypothesis had some inconsistencies /see 2/. The purpose of this review is to discuss the seemingly inconsistent findings concerning the effects of prolonged elevation of glucocorticoids on hippocampal health and to discuss the interpretation that a chronic stress history, which includes repeated elevation of glucocorticoids, may make the hippocampus vulnerable to potential injury
acute stress and the concurrent elevation of glucocorticoids exacerbated damage to the hippocampus caused by neurochemical or metabolic challenges. Glucocorticoids enhance hippocampal damage following neurotoxin insult /kanic acid, also known as 3-acetyl-pyridine, 125,126/, or metabolic challenge /hypoxia, 156/, /hypoglycemia, 156/, /ischemia, 124/. Glucocorticoids can aggravate hippocampal damage when steroid titers are elevated just prior to, during, or immediately after the neurotoxic challenge /122
Down-regulating the HPA axis with repeated glucocorticoid injections to produce low glucocorticoid titers during a hypoxia/ischemia challenge helps to protect against hippocampal damage /72
a damaged hippocampus makes itself even more susceptible to a metabolic challenge than a healthy hippocampus, creating a downward spiral of increased chances of potentiated hippocampal damage with each metabolic episode and this risk increases with age
Only chronic stress, but not acute stress, exacerbated IBO-induced CA3 damage, which confirmed that waiting several days following the end of the last day of restraint permitted glucocorticoid elevations from the last restraint session to subside
Rats exposed repeatedly to the same chronic stress stimulus (homotypic stressor) begin to habituate to that stimulus and show attenuated HPA axis responses. In contrast, rats exposed repeatedly to the same chronic stress stimulus and then presented with a novel stressful stimulus (heterotypic stressor) exhibit potentiated HPA axis responses to the novel stimulus
stress/glucocorticoid-induced hippocampal changes include decreasing neurogenesis /62,108,115/, altering dendritic spine density /97,153,155/, hindering synaptic plasticity /62,115/, impeding long-term potentiation /105/, changing the inhibitory and excitatory tone /62,102,112/, downregulating GR /69,170/, and reducing dendritic complexity /70,82,167
hippocampal CA3 dendritic retraction lasts for at least four days following the end of chronic stress /27,164
chronic stress alters the larger apical region in males as opposed to the smaller basal region in females /49/ with evidence of ovarian hormones being neuroprotective /90/. We found that chronic stress exacerbated IBO-induced CA3 damage in males, but not females /28
dendritic retraction may be an adaptive response to reduce the exposure of CA3 neurons to glucocorticoidinduced glutamate elevation… CA3 dendritic retraction corresponds with impaired spatial ability… neurons that retract dendrites may be unable to respond adequately to an unexpected metabolic challenge, leading to exacerbated hippocampal damage… CA3 dendritic retraction is proposed to be a relatively low-risk, temporary neuroplastic response to combat the immediate looming threat of elevated glutamate
hippocampal dendritic retraction produced by chronic stress in rodents and reduced hippocampal volumes observed in Cushing’s disease, MDD, and PTSD are dynamic and have the capacity to recover… A consistent characteristic of these conditions is that symptom severity correlates with hippocampal size, which shows plasticity by improving with treatment
glucocorticoids appear to be problematic when they are either too high (Cushing’s disease or MDD) or too low (PTSD), but only high levels of glucocorticoids are traditionally thought to reduce hippocampal volumes
animal literature shows that reduced glucocorticoids via adrenalectomy causes neural degeneration within the hippocampal dentate gyrus, /22–24,140/, which can be detected by volumetric measures /23/. Consequently, glucocorticoids can reduce hippocampal volume and plasticity when their levels are either too high or low… inverted U-shaped function
subthreshold metabolic events that are benign to the hippocampus of a healthy individual may be harmful to people with Cushing’s disease, MDD, PTSD or a history of chronic stress
the theory of Cognitive Reserve is used to explain the finding that individuals with more mental activity develop better, more elaborate neuronal connections that can withstand a greater amount of pathological damage before succumbing to disease /66,151/. Individuals engaging in intellectually-stimulating activities throughout their lives maintain their cognitive abilities when others begin to show cognitive decline /1,132,139
housing rats in enriched environments to provide opportunities for social interactions, exercise, and novel visual stimulation prevents the detrimental effects of chronic stress on hippocampal function /171
Copeland W, Keeler G, Angold A, Costello E. 2007. “Traumatic events and posttraumatic stress in childhood.” Arch Gen Psychiat 64:577-584
http://www.ncbi.nlm.nih.gov/pubmed/17485609
Corlew R, Brasier D, Feldman D, Philpot B. 2008. “Presynaptic NMDA receptors: newly appreciated roles in cortical synaptic function and plasticity” Neuroscience 14(6): 609-625
http://nro.sagepub.com/content/14/6/609
Cornelius J, Kirisci L, Reynolds M, Clark D, Hayes J, Tarter R. 2009. “PTSD contributes to teen and young adult cannabis use disorders” Addictive Behaviors 35(2):91-94
http://www.sciencedirect.com/science/article/pii/S0306460309002366
“PTSD contributes to the etiology of CUD among teenagers making the transition to young adulthood beyond the effects of deviant peers, the TLI (Transmissible Liability Index, a measure of risk for SUD), and demographic factors.”
Cornelius J, Aizenstein H, Hariri A. 2010. “Amygdala reactivity is inversely related to level of cannabis use in individuals with comorbid cannabis dependence and major depression” Addict Behav 35(6): 644-646
http://www.ncbi.nlm.nih.gov/pubmed/20189314
a higher level of cannabis use was consistently associated with a lower level of amygdala reactivity across all subjects (matched pairs t=2.70, df=5, p<0.05, two-tailed). These findings are consistent with the reports by Phan et al. (Phan et al., 2008) and Hariri et al. (Hariri et al., 2009) suggesting that cannabinoids have an inhibitory effect on threat-related amygdala reactivity.
major depression and anxiety disorders have … increased reactivity of the amygdala (Drevets 2003), while treatment with antidepressant medications… reduce the reactivity (activation) of the amygdala in subjects with major depression
Cornwall M. 2002. “Alternative Treatment of Psychosis: A Qualitative Study of Jungian Medication-Free Residential Treatment at Diabasis”
https://altmentalities.files.wordpress.com/2012/02/michael-cornwall-diss.pdf
(a) psychosis is not a unitary phenomenon; (b) for at least some people, it is both a purposive developmental process and an experience with strong transpersonal and archetypal elements; and (c) given the right kind of support, such people can come through the experience “healed.”
Participants were strongly critical of the medical model, which Diabasis sought to supplant, but also expressed retrospective criticisms of Diabasis itself
I both observed and facilitated many dramatic recoveries of people in acute psychosis
studies published over the last few decades (e.g. Bola, 1998; Dabrowski & Aronson, 1964; Goldberg, Klerzman, & Cole, 1965; Karon, 1983; Menninger, 1959, 1999; Matthews, Roper, Mosher, & Menn, 1979; Silverman, 1978; Sullivan, 1953) have presented evidence that some people experiencing acute psychotic episodes, when allowed to undergo the psychotic process in a supportive, medication-free setting, can go on to lead rewarding lives, rather than becoming chronic mental patients
for a period of eight years, our therapists were called to the psychiatric emergency screening ward of the hospital whenever somebody without a prior history of psychiatric hospitalization presented with psychotic symptoms. Since we accepted all first break clients, not all of them could have been Brief Reactive
participants treated with medication at Agnews had roughly the same rate of recidivism as the national average in mental hospitals (73%), while the placebo group showed only 8% recidivism
when the Agnews State Hospital Research Department sought to publish the findings, their article was rejected by four major psychiatric journals, despite being approved for publication by the peer review panels assembled by each of the journals. Eventually published in 1978 in International Pharmacopsychiatry, it received little attention in the United States (John Perry, personal communication, January 20, 1998)
Soteria study “has been treated as if it never existed” and described “25 years of swimming upriver against the prevailing biological zeitgeist” (Mosher and Bola, 2000, p. 68)
psychodynamic, existential, humanistic, and transpersonal orientations
Jung (1953, 1960)
for recent annotated bibliographies of studies that demonstrate the brain-damaging effects of neuroleptics, see Mosher (n.d.) and Support Coalition International (1999)
Laing 1967: “true sanity entails, in one way or another, the dissolution of the normal ego, that false self competently adjusted to our alienated social reality; the emergence of the “inner” archetypal mediators of divine power, and through this death a rebirth, and the eventual re-establishment of a new kind of ego-functioning, the ego now being the servant of the divine, no longer its betrayer (p. 145)”
Gestalt Therapy, shamanism, communication theory, and Taoism
Cortes-Briones J, Cahill J, Skosnik P, Mathalon D, Williams A, Sewell R, Roach B, Ford J, Ranganathan M, D’Souza D. 2015. “The psychosis-like effects of Δ9-Tetrahydrocannabinol are associated with increased cortical noise in healthy humans” Biological Psychiatry 78: 805-813
http://www.ncbi.nlm.nih.gov/pubmed/25913109 add to dropbox
The acute, dose-related effects of Δ9-THC on Lempel-Ziv complexity and signal power were studied in humans (n 5 24) who completed 3 test days during which they received intravenous Δ9-THC (placebo, .015 and .03 mg/kg) in a double-blind, randomized, crossover, and counterbalanced design
Δ9-THC increased neural noise in a dose-related manner. Furthermore, there was a strong positive relationship between neural noise and the psychosis-like positive and disorganization symptoms induced by Δ9-THC, which was independent of total signal power. Instead, there was no relationship between noise and negative-like symptoms. In addition, Δ9-THC reduced total signal power during both active drug conditions compared with placebo, but no relationship was detected between signal power and psychosis-like symptoms
Complex mental processes such as perception, language, emotion, and memory rely on the integrity of long-range functional networks formed by ensembles of brain areas (nodes) processing information in a coordinated manner(1 , 2)
converging lines of evidence from structural (diffusion tensor imaging) and functional (functional magnetic resonance imaging [fMRI], electroencephalogram [EEG], and magneto-encephalogram [MEG]) studies have shown abnormal neural connectivity in schizophrenia, which has been related to the presence and intensity of psychotic symptoms ( 3 –9 )
Information is transmitted from one node to another through a medium (e.g., a cable) as part of a signal (e.g., an electromagnetic wave), which is composed by information and random noise. The latter is a sum of random activity and interference caused by other signals travelling through the same medium. According to information theory, the upper limit of the total amount of undistorted (error-free) information per unit of time that can be carried by a signal is limited by: 1) the bandwidth (in Hz), 2) the total power, and 3) the amount of random activity or noise of the signal ( 10–13). Thus, keeping the bandwidth and the total power of a signal constant, the higher the level of noise, the lower the amount of information that can be carried by the signal without distortion. Increased noise may, therefore, disrupt the coordinated activity between nodes, resulting in the disruption of information processing
growing evidence from EEG, MEG, and neuro-imaging studies suggesting increased randomness/noise in the brain activity of schizophrenia patients ( 14–19). A number of studies have shown increased intertrial (random) variability in the latency and amplitude of evoked responses measured in the EEG of schizophrenia patients ( 14, 16–18,20). Furthermore, measures developed to characterize the uncertainty (entropy) or randomness of signals have revealed increased levels of randomness in the EEG and MEG of schizophrenia patients (21–23), which are higher during periods of exacerbation of psychosis (22). While there may be limitations to the existing EEG and MEG literature, recent fMRI data provide further support to the hypothesis that noise is increased in schizo-phrenia ( 19).
schizophrenia patients show increased baseline activity, especially in the gamma band, and an inverse relationship between prestimulation and poststi-mulation gamma activity (43). These findings suggest that at least part of the task-related abnormalities observed in schizophrenia may be associated with the presence of aber-rant brain activity during the prestimulation period
3 test day, randomized, double-blind, placebo-controlled, crossover study, subjects received Δ9-THC (vehicle [ethanol], .015 mg/kg or .03 mg/kg) over 10 minutes by intravenous route. The sample included subjects with and without recent cannabis exposure (within the last 30 days) and excluded cannabis-naïve and cannabis-dependent subjects
there was a significant effect of drug condition on signal power (Wald w22 5 8.004, pAdj 5 .036), which disappeared after controlling for LZC (Wald w22 5 2.236, pAdj ..1). Three HB-adjusted pairwise comparisons performed on the data before controlling for LZC revealed significantly lower power for the .03 mg/kg and .015 mg/kg doses compared with placebo (both pAdj 5 .029) but no difference between the .03 mg/kg and .015 mg/kg doses (pAdj . .1)
regression of LZC on signal power revealed a significant inverse relationship between both variables
significant effect of drug condition for the PANSS positive, disorganization and negative symptoms factors. The nine HB-adjusted pairwise comparisons showed significantly higher scores for the 0.03 mg/kg and 0.015 mg/kg doses compared with placebo and for the .03 mg/kg dose compared with the .015 mg/kg dose
Animal studies have revealed that cannabinoids acutely reduce the spectral power of LFP oscillations by increasing the randomness (reducing the synchronization) of the activity of populations of neurons rather than reducing the activity of individual neurons. The increased randomness would reduce the neurons’ capacity to form temporally coordinated ensembles, leading to a reduction of LFP spectral power (28, 29)
bandwidth of the signals was kept constant (99.5 Hz) across conditions by filtering
In the cerebral cortex and hippocampus, CB1Rs are located on the axon terminals of cholecystokinin-expressing gamma-aminobutyric acidergic interneurons… it has been proposed that cholecystokinin cells enhance the signal-to-noise ratio of neural oscillations (like a noise filter) through a CB1R-mediated mechanism (63–65)
whether our findings apply to resting state activity or to activity associated with expectancy during the prestimulation will need to be determined in future studies
a consequence of using the median as the threshold is that low-range frequencies are preferentially represented in the resulting binary data ( Supplement 1 ). Even though the median-crossing approach is widely used, alternate approaches [e.g., ( 68)] that better represent the entire frequency spectrum should be explored
At doses that produced increases in psychosis-like effects, Δ9-THC increased neural noise (LZC) measured in the EEG of humans in a dose-dependent manner. Furthermore, increases in neural noise were positively related with Δ9-THC-induced psychosis-like, but not negative-like, effects
if psychotic symptoms are a result of brain dysconnectivity related to an abnormal increase of neural noise, interventions directed toward reducing noise may have therapeutic potential
Costello E, Copeland W, Shanahan L, Worthman C, Angold A. 2013. “C-Reactive Protein and substance use disorders in adolescence and early adulthood: a prospective analysis” Drug Alcohol Depend 133(2): 712-717
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4106409/
The acute phase protein C-reactive protein (CRP) is a marker of systemic inflammation (Volanakis, 2001;Black et al., 2004) that is associated with chronic and costly diseases, including cardiovascular and metabolic disease (Raison et al., 2006; Shah et al., 2009; Miller et al., 2011) and with shortened life expectancy (Crimmins and Finch, 2006)
Once established, elevated CRP appears to be fairly stable across time (Macy et al., 1997).
A U-shaped cross-sectional relationship between CRP and alcohol consumption is widely documented (e.g., Albert et al., 2003; Imhof et al., 2001; Pai et al., 2006), but most in studies, including these, alcohol users showing abuse or dependence have elevated CRP
In inflammatory conditions, cannabinoids have long been used to reduce pain in rheumatoid arthritis and Crohn’s disease, and may protect against diabetes (Rajavashisth et al., 2012)
in these analyses we controlled for factors known from the literature to be associated with higher CRP levels (Davey Smith et al., 2005; Benson et al., 2009; Skinner et al., 2010; Elovainio et al., 2006a): age, low socioeconomic status, obesity, psychotropic medications, other medications and some psychiatric disorders
In the simple models without covariates, higher CRP levels at the previous wave predicted a higher likelihood of any cannabis use, but not alcohol use or nicotine use. It predicted abuse or dependence of any drug, and specifically cannabis abuse/dependence and nicotine dependence. When the covariates were added, any abuse/dependence and nicotine dependence remained significant, and there was a trend for cannabis use and abuse/dependence to be predicted by CRP (p<.1)
Only one form of SUD – nicotine use – predicted higher levels of CRP. Significant covariates were age and having a diagnosis of depression
the bivariate cross-sectional associations were much stronger between CRP level and most forms of SUD than they were between CRP and other psychiatric disorders
a recent analysis of the NHANES data (Rajavashisth et al., 2012) found that the prevalence of elevated C reactive protein (>0.5 mg/dl) was significantly less among past or current users than among non-marijuana users, suggesting support for the idea that cannabis is anti-inflammatory
Once other factors were entered into the models, the predictive relationships from CRP to cannabis use and addiction became non-significant
Population studies have consistently shown that there is no clear cut-point between “normal” and elevated levels of CRP (Kushner et al., 2010). In these analyses we excluded values above 10 mg/L (1.5% of all observations) as likely to signal current physical illness. Using the standard definition of “elevated” CRP as >3mg/L, 89.2% of GSMS observations were lower than this, compared with, for example, 62% of about 9,000 subjects (mainly adults) from the 1999–2002 National Health and Nutrition Examination Survey (Woloshin and Schwartz, 2005). Thus, this young and fairly healthy sample may not show effects seen in older or clinical samples.
Cota D. 2008. “The role of the endocannabinoid system in the regulation of hypothalamic-pituitary-adrenal axis activity” J Neuroendocrinol 20 suppl 1:35-8
http://www.ncbi.nlm.nih.gov/pubmed/18426497
previous pharmacological studies using plant-derived or synthetic cannabinoids have implied a stimulating action on the HPA axis … endogenous cannabinoid … inhibiting the release of both adrenocorticotrophic hormone and glucocorticoids
endocannabinoids have been found to mediate the nongenomic glucocorticoid-induced inhibition of the release of corticotrophin-releasing factor within the paraventricular nucleus of the hypothalamus
Cottler L, Nishith P, Compton WM 3rd. 2001. “Gender differences in risk factors for trauma exposure and post-truamatic stress disorder among inner-city drug abusers in and out of treatment” Compr Psychiatry 42(2): 111-7
http://www.ncbi.nlm.nih.gov/pubmed/11244146
Although more women than men met criteria for DSM-III-R PTSD, there were no gender differences on endorsement for a traumatic event. Adult antisocial behavior, affective disorder, schizophrenia, other anxiety disorder and polysubstance use predicted exposure to an event, whereas, only schizophrenia and other anxiety disorder predicted PTSD. In men, drug use preceded the exposure to an event, while in women, the onset age for both drug use and exposure to an event were nearly identical
Cougle J, Bonn-Miller M, Vujanovic A, Zvolensky M, Hawkins K. 2011. “Posttraumatic stress disorder and cannabis use in a nationally representative sample” Psychol Addict Behav 25(3): 554-8
http://www.ncbi.nlm.nih.gov/pubmed/21480682Wh
large representative survey of adults (N = 5,672) from the United States (Kessler et al., 2004)
PTSD diagnoses were associated with increased odds of lifetime history of cannabis use as well as past year daily cannabis use. Lifetime, but not current, PTSD diagnosis also was uniquely associated with increased risk for any past year cannabis use. Additional analyses revealed that the relations between PTSD (lifetime and current) and lifetime cannabis use remained statistically significant when adjusting for co-occurring anxiety and mood disorders and trauma type frequency
Couper K, Blount D, Riley E. 2008. “IL-10: the master regulator of immunity to infection” Journal of Immunology 180(9): 5771-5777
http://www.jimmunol.org/content/180/9/5771.long
IL-10 is an anti-inflammatory cytokine. During infection it inhibits the activity of Th1 cells, NK cells, and macrophages, all of which are required for optimal pathogen clearance but also contribute to tissue damage
IL-10 can act directly on CD4+ T cells, inhibiting proliferation and production of IL-2, IFN-γ, IL-4, IL-5 and TNF-α (5, 8, 9)
Irrespective of the source of IL-10, its effects are similar in all of the infections that have been studied: IL-10 suppresses macrophage and DC function, thereby limiting Th1 and Th2 effector responses (Table I⇑). Nevertheless, the impact of IL-10 is clearly determined by the timing and site of its production, and these are both likely to be affected by which cells are making IL-10. Moreover, because IL-10 production by one cell population can affect the ability of other cells to make IL-10, there is the potential for IL-10-producing cells to regulate each other.
the ability of Th1 cells to coproduce IL-10 and IFN-γ may favor simultaneous pathogen clearance and suppression of downstream pathologies; examples include T. gondii, malaria, and leishmaniasis (17, 37,64, 68)
both Th1 and Th2 cells are now known to mediate inflammation and tissue damage as well as pathogen killing. Anti-inflammatory functions, in contrast, are now known to rest with populations of regulatory cells that may be myeloid or lymphoid in origin and can include fully mature, classical Th1 or Th2 cells that produce IL-10 as a negative feedback mechanism to limit their own response. More than any other cytokine, IL-10 is an essential component of this regulatory response in almost all infections
Cousijn J, Wiers R, Ridderinkhof K, van der Brink W, Veltman D, Goudriaan A. 2012. “Grey mater alterations associated with cannabis use: results of a VBM study in heavy cannabis users and healthy controls.” Neuroimage 59(4): 3845-51
http://www.ncbi.nlm.nih.gov/pubmed/21982932/ in dropbox
Regional grey matter volume in the anterior cerebellum was larger in heavy cannabis users. Within the group of heavy cannabis users, grey matter volume in the amygdala and hippocampus correlated negatively with the amount of cannabis use or dependence. No associations were found between white matter volume and measures of cannabis use or dependence.
Coutinho A, Chapman K. 2011. “The anti-inflammatory and immunosuppressive effects of glucocorticoids, recent developments and mechanistic insights” Mol Cel Endocrinol 335(1): 2-13
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3047790/
Cui M, Yang Y, Yang J, Zhang J, Han H, Ma W, Li H, Mao R Xu L, Hao W, Cao J. 2006. “Enriched environment experience overcomes the memory deficits and depressive-like behavior induced by early life stress”
http://www.ncbi.nlm.nih.gov/pubmed/16790315/
Curran H, Brignell C, Fletcher S, Middleton P, Henry J. 2002. “Cognitive and subjective dose-response effects of acute oral Delta 9-tetrahydrocannabinol (THC) in infrequent cannabis users” Psychopharmacology (Berl) 164(1): 61-70
http://www.ncbi.nlm.nih.gov/pubmed/12373420/
Delta(9)-THC 15 mg impaired performance on two explicit memory tasks at the time of peak plasma concentration (2 h post-drug). At the same time point, performance on an implicit memory task was preserved intact
In several tasks, delta(9)-THC increased both speed and error rates, reflecting “riskier” speed-accuracy trade-offs. Subjective effects were also most marked at 2 h but often persisted longer, with participants rating themselves as “stoned” for 8 h. Participants experienced a strong drug effect, liked this effect and, until 4 h, wanted more oral delta(9)-THC. No effects of delta(9)-THC were found 24 or 48 h following ingestion indicating that the residual effects of oral delta(9)-THC are minimal
Crane N, Schuster R, Fusar-Poli P, Gonzalez R. 2012. “Effects of cannabis on neurocognitive functioning: recent advances, neurodevelopmental influences, and sex differences” Neuropsychol Rev 23:117-137
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3593817/
the neurobehavioral effects of cannabis use have been intensely studied over several decades and summarized in numerous published reviews (Crean et al. 2011; Ferraro 1980; R. Gonzalez et al. 2002; R. Gonzalez 2007; I. Grant et al. 2003; Iversen 2003; Jager and Ramsey 2008; Pope et al. 1995; Ranganathan and D’Souza 2006; Schweinsburg et al. 2008; Solowij and Battisti 2008; Solowij and Pesa 2010; Wert and Raulin 1986). However, many of these reviews conclude that the findings for several neurocognitive domains are equivocal
Several review articles have been written on acute cannabis intoxication and neuropsychological functioning (Crean et al. 2011; R. Gonzalez 2007; Ferraro 1980;Ranganathan and D’Souza 2006; Solowij and Pesa 2010), with equivocal findings for most neurocognitive domains other than aspects of episodic memory – the autobiographical memory of specific events, situations, and experiences (Tulving 2001). … More recent studies also suggest that acute THC intoxication seems to impair retrieval-based memory of new material, but evidence regarding the effects of THC on other neuropsychological domains remained mixed (R. Gonzalez 2007; Ranganathan and D’Souza 2006). For instance, a recent review of cannabis’ effects on executive functioning suggests that THC administration adversely affects inhibition, impulsivity, and working memory, but not verbal fluency, and findings are mixed for decision-making, risk-taking, and aspects of attention (Crean et al. 2011)
Data from the past five years is consistent with previous studies, generally showing acute administration of THC to impair episodic memory, including immediate (D’Souza et al. 2008a; D’Souza et al. 2008b; Dumont et al. 2011;Morrison et al. 2009) and delayed recall (D’Souza et al. 2008a; D’Souza et al. 2008b); procedural memory (Dumont et al. 2011); and associative learning and memory (Ballard et al. 2012) among occasional and regular cannabis users as well as non-users. Evidence indicates there may be a dose-dependent relationship between amount of intravenous THC and impairments in total free and delayed verbal recall, but not delayed cued or recognition recall (D’Souza et al. 2008b)
In studies of visuospatial memory, THC impaired accuracy in regular cannabis users (Weinstein et al. 2008a; Weinstein et al. 2008b). On the other hand, others report no effect from THC on visuospatial memory (Anderson et al. 2010), associative memory (Bossong et al. 2012a), or verbal episodic memory (Sugarman et al. 2011). Hart and colleagues (2010) also found that THC administration did not affect overall performance on a task of verbal episodic memory in regular cannabis users, but participants performed more poorly on a recognition task suggesting a deleterious impact of cannabis on retrieval-based memory
Most studies document impairments in attention and concentration following administration of small (i.e., 2.5mg) and large (i.e., 0.5 mg/kg) doses of THC in cannabis users and non-users compared to placebo administration (Anderson et al. 2010; D’Souza et al. 2008a; D’Souza et al. 2008b; Hunault et al. 2009; Ramaekers et al. 2009; Ramaekers et al. 2011;Theunissen et al. 2011), with some evidence for a dose-dependent relationship between amount of THC smoked and degree of impairment (Hunault et al. 2009). However, two studies found no differences in performance on measures of auditory selective attention and concentration in regular cannabis users after smoking cannabis standardized to 20mg THC (O’Leary et al. 2007) or on a measure of sustained attention in occasional cannabis users after ingesting 15mg THC (Sugarman et al. 2011) compared to placebo. Impairments in working memory have been observed following acute administration of vaporized THC (Bossong et al. 2012b), oral THC (D’Souza et al. 2008a; Dumont et al. 2011), smoked cannabis containing different doses of THC (Hart et al. 2010), and intravenous THC (Morrison et al. 2009)
THC has been found to impair healthy young adults’ performance on a reasoning task (but not a verbal fluency task; Morrison et al. 2009) and significantly increased total errors and nonpreservative errors in regular cannabis users on a task of abstract reasoning (Weinstein et al. 2008a; Weinstein et al. 2008b). Additionally, problems with cognitive flexibility have been reported after smoking cannabis standardized to different doses of THC compared to a placebo condition (Anderson et al. 2010). In contrast, most current evidence suggests that acute intoxication does not negatively impact decision-making or risk-taking in occasional, regular, or heavy cannabis users (D’Souza et al. 2008a; Ramaekers et al. 2009; Ramaekers et al. 2011; Vadhan et al. 2007; Weinstein et al. 2008a; Weinstein et al. 2008b), but it may slow decision-making (Vadhan et al. 2007). Some contradictory evidence does exist. Whereas a previous study demonstrated increasing levels of cannabis intoxication were associated with more risky responses in occasional cannabis users (Lane et al. 2005), a recent study found THC may actually reduce risk-taking behaviors in healthy young adults (Rogers et al. 2007)
Similar to the findings for decision-making and risk taking, studies examining inhibition, motor impulsivity, and psychomotor control (these domains are invariably linked and generally defined as behavior involving rapid reactions to internal and external stimuli; Moeller et al. 2001) are mixed. THC was found to negatively influence inhibitory control, as evidenced by increased stop reaction time and decreased accuracy of responses in occasional and heavy cannabis users during a stop signal task in two studies (Ramaekers et al. 2009; Theunissen et al. 2011), but THC administration, three hours after placebo alcohol administration, did not have an effect on inhibition or motor impulsivity in heavy cannabis users on the same task in another study (Ramaekers et al. 2011). Given the motor components inherent in the aforementioned tasks, it is worth considering that some studies find THC to impair psychomotor performance (D’Souza et al. 2008a; Hunault et al. 2009; Ramaekers et al. 2009), but several others do not (Dumont et al. 2011; O’Leary et al. 2007; Ramaekers et al. 2011). Interestingly, Ramaekers and colleagues (2009) found THC administration impaired psychomotor control in occasional cannabis users, but not in heavy cannabis users, suggesting a potential tolerance effect.Hunault and colleagues (2009) found THC to significantly decrease response time and increase errors in a dose-dependent manner in heavy cannabis users on a motor control task
Borgwardt and colleagues (2008) administered 10mg THC, 600 mg CBD, or placebo capsules to 15 men with minimal cannabis exposure (<15 times in life) on three separate occasions in a double-blind, within-subject design. In this study, and across several other investigations using the same sample, participants’ performance was unaffected by THC or CBD on measures of inhibition, verbal memory, accuracy of gender discrimination, and visual and auditory processing (Bhattacharyya et al. 2009; Bhattacharyya et al. 2010; Borgwardt et al. 2008; Fusar-Poli et al. 2009; Winton-Brown et al. 2011). In contrast, Roser and colleagues (2009) found combined THC and CBD, but not THC alone, impaired psychomotor control in 24 occasional cannabis users who were administered a combination of 10mg THC and 5.4mg CBD, 10mg THC alone, or a placebo pill on three separate visits in a double-blind, cross-over design
Morgan and colleagues (2010)found 22 cannabis users with low cumulative levels of CBD (as evidenced from hair analysis) demonstrated poorer performance on immediate and delayed episodic memory (but not verbal and category fluency) compared to 22 users with higher cumulative CBD exposure when both groups were intoxicated. The same group showed that daily cannabis users who had higher cumulative THC exposure demonstrated poorer performance on measures of immediate and delayed episodic memory, as well as source memory, compared to daily cannabis users with lower cumulative THC exposure when participants were intoxicated (Morgan et al. 2011). However, there were no differences in episodic memory and source memory performance between the 54 occasional cannabis users regardless of the cumulative THC exposure. On the other hand, both daily and occasional cannabis users with CBD present in hair analysis displayed better recognition memory than users without CBD present when all individuals were intoxicated (Morgan et al. 2011).
although no neurocognitive differences were found, THC and CBD had opposite effects of brain activation patterns in the striatum during verbal recall, in the hippocampus during inhibition, in the superior temporal cortex during auditory processing, in the occipital cortex during visual processing, and in the amygdala while subjects viewed fearful faces when compared to placebo (Bhattacharyya et al. 2010).
Some suggest residual effects (i.e., after several hours to several days of abstinence) and long-term effects (i.e., after several weeks of abstinence and beyond), while others find no clear evidence for residual or long-term neuropsychological deficits among cannabis users across several domains (Crean et al. 2011; R. Gonzalez et al. 2002; R. Gonzalez 2007; I. Grant et al. 2003; Pope et al. 1995; Schweinsburg et al. 2008; Solowij and Battisti 2008; Solowij and Pesa 2010; Wert and Raulin 1986). As found in the acute intoxication studies, the evidence is mixed for most domains other than episodic memory, which remains adversely impacted for up to 28 days following abstinence (R. Gonzalez 2007; I. Grant et al. 2003; Solowij and Battisti 2008)
Regular adolescent cannabis users who have been abstinent between 12 hours and 21 days demonstrate poorer immediate (Hanson et al. 2010;Harvey et al. 2007; Solowij et al. 2011) and delayed recall (Harvey et al. 2007; Solowij et al. 2011), as well as impaired recognition (Solowij et al. 2011). However, one of these studies found no difference in recognition between regular and occasional cannabis users (Harvey et al. 2007). After longer periods of abstinence (28 days), Tapert and colleagues (2007) found adolescent cannabis users made more recall intrusions than controls, while other studies found no differences between groups on any measure of episodic memory (Jacobsen et al. 2007; Mahmood et al. 2010; Medina et al. 2007a). Recently abstinent adult cannabis users also demonstrate poorer immediate (Battisti et al. 2010b; R. Gonzalez et al. In press;Hadjiefthyvoulou et al. 2011; Korver et al. 2010; Nestor et al. 2008; Wagner et al. 2010; Yucel et al. 2008) and delayed recall (R. Gonzalez et al. In press; Wadsworth et al. 2006; Yucel et al. 2008), but have intact recognition (R. Gonzalez et al. In press; Nestor et al. 2008; Wadsworth et al. 2006). Although this is not invariably the case, as some studies report intact immediate (Chang et al. 2006; Gruber et al. 2011;Wadsworth et al. 2006) and delayed recall (Chang et al. 2006; Gruber et al. 2011) among cannabis users. Other evidence suggests that recall performance is negatively associated with amount of past year (Jager et al. 2007) and lifetime cannabis use (Indlekofer et al. 2009; Jager et al. 2007; P. Murphy et al. 2011;Solowij et al. 2011), duration of cannabis use (Solowij et al. 2011; Wadsworth et al. 2006), frequency of cannabis use, and age of first cannabis use (Solowij et al. 2011).
Adolescent cannabis users who were abstinent between 12 to 24 hours demonstrated intact associative learning (Harvey et al. 2007; Jager et al. 2010). After longer periods of abstinence (28 days), adolescent cannabis users exhibited deficits on a story memory task (Medina et al. 2007a), but had intact visuospatial memory (Mahmood et al. 2010; Medina et al. 2007a; Schweinsburg et al. 2010). In recently abstinent adult cannabis users, some studies report visuospatial memory deficits (Hermann et al. 2007; McHale and Hunt 2008), but others do not (Chang et al. 2006; Gruber et al. 2011). Amount of lifetime cannabis use did not predict visuospatial memory in a community sample of recently abstinent polydrug users (Indlekofer et al. 2009). Recently abstinent adult cannabis users have demonstrated intact associative learning (Fisk and Montgomery 2008) and semantic memory (Wadsworth et al. 2006). Evidence for prospective memory deficits is split with one study reporting deficits (Montgomery et al. 2012) and another no deficits (Hadjiefthyvoulou et al. 2011)
Several studies cite impairments in attention and concentration in recently abstinent adolescent cannabis users (Abdullaev et al. 2010; Hanson et al. 2010; Harvey et al. 2007; Lane et al. 2007). After at least 21 days of abstinence, complex attention continues to be impaired in adolescent cannabis users (Hanson et al. 2010; Jacobsen et al. 2004; Medina et al. 2007a; Tapert et al. 2007), with evidence of a dose-dependent relationship with amount of lifetime use (Medina et al. 2007a). Conversely, others reported that after 45 days of abstinence, selective and divided attention was intact in adolescent cannabis users (Jacobsen et al. 2004). Recently abstinent adult cannabis users also demonstrated impairments in attention and concentration in several studies (Hermann et al. 2007; Indlekofer et al. 2009;Scholes and Martin-Iverson 2009; Scholes-Balog and Martin-Iverson 2011; Wadsworth et al. 2006), which persisted for several hours to one week of abstinence. However, a few studies report no impairments among adult cannabis users when the time of last cannabis use was either unknown (J. Grant et al. 2011;Korver et al. 2010) or after at least 4 hours to approximately 38 months of abstinence (Chang et al. 2006)…
Adolescent regular cannabis users who are abstinent demonstrated poorer working memory compared to controls after 3 and 13 days, during a challenging task that required active manipulation of items (Hanson et al. 2010), but not after 8 days, during a more simple matching task (Jager et al. 2010) … Harvey and colleagues (2007) found regular adolescent cannabis users performed more poorly on spatial working memory than occasional cannabis users and amount of cannabis use was one of the strongest predictors of poorer spatial working memory. The evidence is also mixed regarding the persistence of working memory deficits among adolescents after abstinence, with some studies reporting deficits after 28 days (Jacobsen et al. 2004; Jacobsen et al. 2007), whereas others do not (Hanson et al. 2010; Padula et al. 2007; Schweinsburg et al. 2008; Schweinsburg et al. 2010). In contrast to studies with adolescent samples, most studies report intact working memory among recently abstinent adult cannabis users (Becker et al. 2010; Chang et al. 2006; Fisk and Montgomery 2008; J. Grant et al. 2011; Gruber et al. 2011; Jager et al. 2006; Looby and Earleywine 2010; Piechatzek et al. 2009; Scholes and Martin-Iverson 2010)… in a longitudinal study, working memory performance remained intact among recently abstinent current cannabis users, former cannabis users, and controls over a period of eight years (Tait et al. 2011). On the other hand, frequency of cannabis use was negatively correlated with working memory performance among abstinent cannabis users (Wadsworth et al. 2006) and among polysubstance users (Fernandez-Serrano et al. 2010)
one recent study examined verbal fluency in a sample of adolescent cannabis users and found it to be impaired after 28 days of abstinence (Tapert et al. 2007). Studies of recently abstinent adult cannabis users report deficits in verbal fluency after 8-24 hours since last use (Korver et al. 2010; Mason et al. 2012;McHale and Hunt 2008) and after 15 days of abstinence (Fernandez-Serrano et al. 2010). However, others report intact verbal fluency after a few hours to a week since last cannabis use (Chang et al. 2006; Fisk and Montgomery 2008; Gruber et al. 2011; Piechatzek et al. 2009). Of note, recently abstinent adult cannabis users have also been reported to demonstrate deficits in text comprehension (Huestegge et al. 2010)
Poorer cognitive flexibility has been reported among adolescent cannabis users abstinent for approximately 8 hours compared to controls (Lane et al. 2007), but not between recently abstinent occasional and regular adolescent cannabis users (Harvey et al. 2007). After at least 23 days of abstinence, adolescent cannabis users appear to evidence intact cognitive flexibility (Medina et al. 2007a). As with adolescent cannabis users, abstinent adult cannabis users also demonstrated impairments in cognitive flexibility (Fontes et al. 2011a, 2011b; J. Grant et al. 2011; Mason et al. 2012). Similarly, planning and reasoning were poorer among adult cannabis users who were abstinent for at least 5 days compared to controls (Montgomery et al. 2012). Furthermore, amount of lifetime cannabis use predicted worse performance on a reasoning task for adult polysubstance users who were abstinent for at least 15 days (Fernandez-Serrano et al. 2010)
Compared to findings from acute investigations, recently abstinent adult cannabis users show more consistent deficits in decision-making and risk-taking (J. Grant et al. 2011;Vaidya et al. 2012; Wesley et al. 2011), even after 15 days of abstinence (Fernandez-Serrano et al. 2010).Fridberg and colleagues (2010) also reported impairments in decision-making among cannabis users, and through using computational modeling they found these impairments were likely due to cannabis users’ decreased sensitivity to losses and increased sensitivity to immediate gains. However, others have not observed such deficits (Boggio et al. 2010; Hermann et al. 2009; Martin-Soelch et al. 2009). In our own studies, cannabis users and non-users, who were matched on various potential comorbidities, performed equivalently on measures of decision-making and risk-taking (R. Gonzalez et al. In press). However, we found that decision-making performance (but not performance on measures of episodic memory or other measures associated with impulsive behavior) is related to more DSM-IV cannabis use dependence symptoms among cannabis users (R. Gonzalez et al. In press)
recently abstinent adolescent cannabis users, regular and non-regular users showed no difference in inhibition or psychomotor control (Harvey et al. 2007). After 28 days of abstinence, adolescent cannabis users demonstrated intact inhibition and motor impulsivity (Tapert et al. 2007). Yet, after at least 23 days of abstinence, adolescent cannabis users had impairments in psychomotor performance compared to controls, and there was a negative, dose-dependent association between performance and lifetime cannabis use episodes (Medina et al. 2007a). In recently abstinent adult cannabis users, some studies report impairments in inhibition and motor impulsivity (Battisti et al. 2010a; Clark et al. 2009; Cunha et al. 2010; Fontes et al. 2011a, 2011b; Scholes and Martin-Iverson 2010), with some evidence for a dose-dependent relationship between amount of past 30 day use and performance (Cunha et al. 2010; Piechatzek et al. 2009). However, many other studies report no differences in performance between recently abstinent adult cannabis users and controls (Aharonovich et al. 2008; Becker et al. 2010;Cane et al. 2009; Chang et al. 2006; Fernandez-Serrano et al. 2010; Fontes et al. 2011b; R. Gonzalez et al. In press; J. Grant et al. 2011; Gruber et al. 2011; Hermann et al. 2007; Hester et al. 2009; Roberts and Garavan 2010; Scholes and Martin-Iverson 2010)
one investigation finding deficits in psychomotor control across several measures among adult cannabis users abstinent for 12 hours (King et al. 2011), but others suggesting intact psychomotor control in recently abstinent cannabis users (Chang et al. 2006; Korver et al. 2010; Wadsworth et al. 2006) and after 28 days of abstinence (Pillay et al. 2008)
there is some evidence indicating cannabis may exert neuroprotective effects among heavy alcohol using adolescents (Jacobus et al. 2009; Mahmood et al. 2010; Medina et al. 2007b; Schweinsburg et al. 2011), alcohol-dependent adults (Nixon et al. 1998), and methamphetamine-dependent adults (R. Gonzalez et al. 2004)
rats exposed to synthetic cannabinoids or THC during adolescence experience impaired working memory during adulthood (O’Shea et al. 2004; O’Shea et al. 2006; Rubino et al. 2009a; Rubino et al. 2009b). These impairments have been correlated with less active synapses in the prefrontal cortex (Rubino et al. 2009a), as well as shorter dendrites and reduced spine densities in the hippocampus, suggesting enduring neurobiological consequences of early cannabis exposure (Rubino et al. 2009b). The heightened vulnerability of younger brains is highlighted by the fact that the same amount of THC exposure that led to decreased working memory performance in adolescent rats had no effect in adult rats (Quinn et al. 2008). However, some have suggested that heavy THC administration during adolescence may be needed to produce significant and persistent impairments in working memory (Realini et al. 2009). With regards to learning, impairments with synthetic cannabinoid administration during late adolescence were reported, but these resolved after a prolonged period of abstinence (Abush and Akirav 2012). Conversely, rats exposed to chronic doses of THC during adolescence, but not during late adolescence, evidenced deficits in learning during adulthood (Harte and Dow-Edwards 2010). There may be a critical time during adolescence when cannabis use may have the most negative effects. When taken together, these studies suggest that cannabis use during adolescence may have a more significant and lasting impact on neurocognition than if cannabis use is initiated during adulthood.
those who initiate use before 15 to 17 years of age demonstrate more pronounced deficits in visual attention (Ehrenreich et al. 1999), verbal fluency (Gruber et al. 2011; Pope et al. 2003), inhibition (Fontes et al. 2011a; Gruber et al. 2011; Pope et al. 2003), and other aspects of executive functioning (Fontes et al. 2011a) as compared to those who initiate use later on. Further, episodic memory was poorer in those with an earlier onset of cannabis use in two studies (Pope et al. 2003; Solowij et al. 2011); however, another study failed to find this effect (Gruber et al. 2011). Poorer performance on measures of inhibition (Battisti et al. 2010a; Gruber et al. 2011) and impulsivity (Solowij et al. 2012) have also been associated with earlier age of onset. Moreover, Gruber and colleagues (2012) reported that early-onset users made more errors and showed greater disruptions in brain activation patterns than late-onset users during an inhibition task. On the other hand, others have reported no differences between early and late-onset cannabis users (and healthy controls) on measures of working memory and attention (Ehrenreich et al. 1999; Gruber et al. 2011; Pope et al. 2003), as well as on a task of visuospatial memory (Gruber et al. 2011)
Females’ total brain size peaks between 10 and 11 years of age, while males’ total brain size peaks at about 14 to 15 years of age (Lenroot et al. 2007). Similarly, females’ prefrontal cortex gray matter volume peaks 1 to 2 years earlier than in males (Giedd et al. 1999). Therefore, if cannabis use is initiated in early to mid-adolescence, it is possible that males may be more vulnerable to neurobehavioral disturbances, especially functional and/or structural disruptions in the prefrontal cortex, compared to females
nimal studies citing greater CB1 receptor density among males across several brain regions (Burston et al. 2010; Mateos et al. 2011; Reich et al. 2009; Rubino et al. 2008) Whereas males either maintain or lose some CB1 binding sites in later adulthood (i.e., 45 to 70 years of age), adult female brains show increases in CB1 receptor density across the lifespan, eventually surpassing that of males (Van Laere et al. 2008)
Furthermore, adolescent females evidence greater CB1 desensitization after exposure to THC in the prefrontal cortex, hippocampus, striatum, amygdala, and midbrain (Burston et al. 2010; Rubino et al. 2008). Given that endocannabinoid signaling plays a crucial role in establishing normal sex differences in the brain (Viveros et al. 2012)
Acute THC administration disrupted learning in adolescent female, adolescent male, and adult female rats, but did not disrupt learning in adult male rats (Cha et al. 2007). Chronic doses of THC administration did not affect learning in either adolescent or adult male and female rats (Cha et al. 2007), but others found that male rats evidenced worse novel object recognition than female rats (Mateos et al. 2011). In contrast, female rats administered chronic THC during adolescence perform worse on an object location task than female controls, while no differences emerged among male rats (Mateos et al. 2011). Although both male and female rats demonstrated deficits in spatial working memory after exposure to THC in adolescence (O’Shea et al. 2004; O’Shea et al. 2006; Rubino et al. 2009a; Rubino et al. 2009b), this exposure in adulthood had long-term impairments on memory in male rats, but not in female rats (O’Shea et al. 2004; O’Shea et al. 2006). Thus, cannabis use during adolescence may produce sex-specific alterations in neurodevelopment
evidence from animal studies suggest exogenous cannabinoids, like THC, have the ability to significantly alter pituitary function (L. Murphy et al. 1991a; L. Murphy et al. 1991b; Wenger et al. 1999) and may inhibit the HPG axis (L. Murphy et al. 1990; L. Murphy et al. 1994;L. Murphy et al. 1999; Steger et al. 1980; Steger et al. 1983), thereby dysregulating key motivational systems involved in drug-seeking behavior
estrogen reduces the inhibitory effect of cannabinoids on HPG function (Scorticati et al. 2003; Scorticati et al. 2004). Furthermore, hormones play a key role in structural and functional changes in the endocannabinoid system, evidenced by fluctuations in endocannabinoid receptor (especially CB1) density across the estrus cycle (Bradshaw et al. 2006; S. Gonzalez et al. 2000; Rodriguez de Fonseca et al. 1994) Moreover, human studies have demonstrated that, during the follicular phase of the menstrual cycle (when estradiol is high and progesterone is low), females are more responsive to the effects of stimulants, such as cocaine (Sofuoglu et al. 1999) and nicotine (Newman 2009; Perkins et al. 2000)
females enter treatment for marijuana use disorders significantly sooner (i.e., after fewer years of use) and after less cumulative cannabis use compared to males (Hernandez-Avila et al. 2004). This may be due to various differences in the pharmacologic effects of THC in males and females
female rats preferentially metabolize THC to its most highly active metabolite, while male rats metabolize THC to multiple compounds (Narimatsu et al. 1991). Moreover, physiological data shows that when smoking a second cigarette, females have less tachycardia than males (Cocchetto et al. 1981)
females may feel greater hedonic reinforcement from cannabis compared to males, and that ovarian hormones (such as estrogen and progesterone) may facilitate stronger learned associations between drug effects and drug-related stimuli (Fattore et al. 2007)
it is possible that females may be more vulnerable to the negative effects of cannabis
Males often perform better than females on measures of decision-making (Bolla et al. 2004; Overman et al. 2004;Reavis and Overman 2001), spatial processing, and psychomotor performance (Gur et al. 2012); whereas females perform better on measures of verbal memory (Kramer et al. 1988), visual recognition, attention, and reasoning abilities (Gur et al. 2012)
35 male and 35 female cannabis users did not demonstrate differential effects of smoked THC (joints with either 0% THC or 2.9% THC) in performance on measures of visuospatial processing, time estimation, and cognitive flexibility (Anderson et al. 2010)
Rogers and colleagues (2007) found no sex differences in risk-taking performance between 7 male and 8 female cannabis users after administration of either 0mg or 5mg THC. Surprisingly, spatial working memory was enhanced in females (n= 7), but not in males (n=12) after 0mg or 8.5mg of sublingual THC (Makela et al. 2006)
in all five sessions of a finger tapping task, males (n= 12) had faster left-hand tapping than females (n= 12) after 10mg THC administration and for two sessions after THC and CBD combined administration (10mg THC and 5.4mg CBD), despite no differences in baseline performance (Roser et al. 2009). In summary, preliminary evidence suggests there may be little or no sex-specific effects of acute THC administration on neurocognition
females who used cannabis heavily (i.e., median frequency of use was 29 out of the past 30 days) performed worse than light users (i.e., median frequency of use was 1 out of the past 30 days) on a visuospatial task – no differences were found between heavy and light male cannabis users, nor were any sex differences found on task of attention and inhibition (Pope et al. 1997). More recently, male cannabis users evidenced impairments on psychomotor performance (Clark et al. 2009; King et al. 2011) and visuospatial organization (King et al. 2011) compared to male healthy controls, but no differences were observed between cannabis-using females and their non-using counterparts (Clark et al. 2009; King et al. 2011) among male (n= 44) and female (n = 25) young adult cannabis users (unpublished). Specifically, more lifetime cannabis use was associated with poorer decision-making performance for male cannabis users, but not female cannabis users. In contrast, more lifetime cannabis use was negatively associated with episodic memory performance in both male and female cannabis users. Other studies report no interaction effects of sex and cannabis use on episodic memory (Solowij et al. 2011; Tait et al. 2011) or working memory (Tait et al. 2011)
After 28 days of abstinence, female adolescent chronic cannabis users had larger right amygdala volumes compared to female adolescent healthy controls, while there was no difference in amygdala volumes among adolescent male chronic cannabis users and controls (McQueeny et al. 2011). Female adolescent chronic cannabis users also evidenced larger prefrontal cortex volumes than female controls, whereas males showed the opposite pattern relative to a male control group (Medina et al. 2009). In this sample, smaller prefrontal cortex volume was associated with better executive functioning among the cannabis users, but larger prefrontal cortex volume was associated with better executive functioning among the controls
We suggest that a specific focus on pre-existing neurobehavioral factors and sex-differences, as well as their complex interactions, will be fruitful in understanding the subjective liability to the neurocognitive effects of cannabis.
Crippa J, Zuardi A, Martin-Santos R, Bhattacharyya S, Atakan Z, McGuire P, Fusar-Poli P. 2009. “Cannabis and anxiety: a critical review of the evidence” Hum. Psychopharmacol Clin Exp 24: 515-523
http://www.ncbi.nlm.nih.gov/pubmed/19693792 add to dropbox
Cannabis can cause an acute and short-lasting episode of anxiety, which often resembles a panic attack, in those who are not habitual users… About 20-30% of users show brief acute anxiety reactions after smoking the drug (Hollister, 1986; Thomas, 1996). Such symptoms usually occur when the drug is used at high doses aand are more common in drug-naive subjects and in novel or stressful environmental situations (Manzanares et al, 2004)
consistently described in experimental studies in man (D’Souza et al ., 2004; Gregg et al ., 1976; Naliboff et al ., 1976; Zuardi et al ., 1982) and in many case reports (Deaset al ., 2000; Moran, 1986)
anxiety reactions and panic attacks appear to result in aversion to further use of the drug, as these symptoms are one of the most common reasons given for the cessation of cannabis use. These individuals are also less likely to be come regular users (Thomas, 1996)
cannabis can exacerbate existing anxiety symptoms, precipitate recurence of an acute episode and counteract the effects of anxiolytic medication (Szuster et al, 1988)
Factors associated with the risk of anxiety induced by cannabis use: Individual and genetic vulnerability Personality traits Gender Frequency of use Dose and quantity consumed Proportions and concentration of cannabinoids History of previous episode Presence of anxiety disorders/symptoms Basal anxiety levels Abstinence states Environment and context of use
In contrast, long-term users typically report that cannabis use is associated with a reduction in anxiety. Relaxation and relief from tension remain the most common effects of using cannabis and the most common reasons for using the drug (Boys et al, 1999; Hathaway, 2003; Reilly et al, 1998).
Frequent cannabis users appear to have higher levels of anxiety than non-users without these necessarily representing an anxiety disorder. In a study with people who regularly used cannabis for at least 10 years, 21% of the subjects had high levels of anxiety (Reilly et al, 1998)
Troisi: severity of anxiety symptoms increased progressively with increasing cannabis use and higher levels of anxiety were observed in those who reported abuse or dependence on the drug
Dorard (2008) confirmed that cannabis dependence in adolescents is related to increased psychological distress and anxiety levels. Similar result was found in two indigenous communities in the northern territory of Australia (Clough et al., 2005). The authors found that the severity of symptoms within the ‘anxiety-dependency’ cluster in cannabis users increased as their level of cannabis use increased
particularly evident among women, as observed in cannabis-dependent individuals arrested for marijuana-related offending behaviour (Feeney et al ., 2005). State anxiety level was also one of the main predictors of increased severity of cannabis use (Spalletta et al ., 2007), while the marijuana use increased the risk for anxiet y symptoms and catastrophic thinking (Zvolensky et al ., 2006b)
a considerable number of subjects developed anxiety disorders before the first symptoms of cannabis dependence suggesting that some of them may have been using cannabis as a self-prescribed anxiolytic medication (Agosti et al., 2002). In line with this hypothesis a recent study observed that social anxiety disorder is an independent risk factor for cannabis dependence (Buckner et al ., 2008)
authors of this study found that the age of onset of panic attacks was significantly earlier among the subjects with both conditions than for individuals with panic attack with no cannabis use (Zvolensky et al ., 2006a)
Strike et al . (2003) verified that 13% of individuals dependent on cannabis who sought treatment had a history of previous anxiety disorder. Similarly, Swadi and Bobier (2003) verified that 63% of patients hospitalized with an anxiety disorder presented with a comorbid substance abuse disorder, especially cannabis. On the other hand, suffering from an anxiety disorder seems to be one of the most important reasons for cannabis users to seek psychiatric treatment
An-xiety is one of the main manifestations of the cannabis withdrawal syndrome (Bonn-Miller et al., 2007; Haney, 2005). Its onset is typically between the second and sixth day and lasts from 4 to 14 days (Budney et al ., 2004). The severity and duration of these effects are comparable to those of tobacco and other withdrawal syndromes… The most promising demonstration of suppression of symptoms of cannabis withdrawal was observed through the use of oral D9-THC (from 30 to 90 mg/day), which decreased both symptoms and cravings. At this dose
it did not produce intoxication and induced only minimal adverse effects (Budney et al ., 2007; Haney et al ., 2004)
[MV: 90 mg THC is a LOT… “did not produce intoxication”?]
Several cases of agoraphobia (Moran, 1986) and panic disorder (Dannon et al ., 2004; Langs et al ., 1997) induced by cannabis use have been previously described. Some authors have also speculated that cannabis use may be associated with enduring anxiety symptoms, as these clinical observations parallel data from animal studies (O’Shea et al ., 2004, 2006).
It was observed that frequent cannabis use in adolescent females doubled the chances of their subsequently presenting with anxiety and depressive conditions, with daily cannabis use increasing the risk fourfold (Patton et al ., 2002). As depression did not predict higher cannabis use in later life, this seems unlikely to reflect self-medication with cannabis for these symptoms; consistent with an earlier report (McGee et al., 2000). These findings have been replicated by a recent study which followed up a cohort of 3239 Australian young adults from birth to the age of 21 years. After controlling for confounding factors, those who started using cannabis before the age of 15 years and used it frequently at 21 years were more likely to report symptoms of an anxiety disorder in early adulthood (Hayatbakhsh et al., 2007). However, contrasting results were observed in a large longitudinal study that focused on the vulnerability factors in adolescent substance use (Windle and Wiesner, 2004). These contrasting findings were further supported by a recent study which investigated whether cannabis use predicted the first incidence of mood and anxiety disorders in 6000 adults during a 3-year follow-up period. Use of cannabis at baseline predicted a modest increase in the risk of a first episode of major depression and a stronger increase in the risk of a first episode of bipolar disorder. Although there was an association noted between cannabis use and anxiety disorders, it did not survive after controlling for confounding factors (Van Laaret al ., 2007)
the combination of cognitive-behavioural therapy and pharmacotherapy was not effective for the treatment of panic or social phobia symptoms among occasional cannabis users (Brickeret al ., 2007)
a causal relationship between cannabis use and long-term anxiety disorders remains dubious (Gilder et al., 2006; Windle and Wiesner, 2004). For instance, if cannabis use were to have been responsible for the precipitation of anxiety disorders, one would have expected an increase in the number of people presenting with these disorders in recent years, as the prevalence and extent of cannabis use has increased in recent times (Boydell et al ., 2006; Iversen, 2003)
Cuthbert B, Insel T. 2013. “Toward the future of psychiatric diagnosis: the seven pillars of RDoC” BMC Med 11: 126
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3653747/
Current diagnostic systems for mental disorders rely upon presenting signs and symptoms, with the result that current definitions do not adequately reflect relevant neurobiological and behavioral systems – impeding not only research on etiology and pathophysiology but also the development of new treatments
Dallman M. 2007. “Modulation of stress responses: how do we cope with excess glucocorticoids” Exp Neurol 206(2): 179-182
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2795792/
Circulating glucose is increased with an abundant supply for the potential need for fighting or fleeing, as is arterial blood pressure; the immune system is prevented from over-responding to acute bacterial threats; contextual brain-generated behaviors are enhanced by both increased attention and drive (13). Thus, the glucocorticoids engage many of the body systems for optimal responses to acute challenge
persistently elevated glucocorticoid concentrations are far too much of a once good thing
there appears to be a host of mechanisms that allow strong damping of the HPA responses
“habituation” to (homotypic) stressors … with repetition the HPA response diminishes markedly, thus limiting the overall exposure to glucocorticoids experienced by the animal during repeated bouts of the same stimulus… the HPA system maintains, or even augments its responsivity to new (heterotypic) stimuli
all indices of activity are diminished after repetition. Corticotropin-releasing factor (CRF) and vasopressin(AVP) synthesis is decreased as is consequent ACTH and corticosterone secretion (6) … c-Fos staining, shows a reduced number of excited cells, and that the responsive CRF neurons are more ventrally located in the nuclei (20)
neurotoxin lesions of the posterior midline paraventricular thalamus, known as the neuroendocrine thalamus, abrogated habituation to repeated stressors… habituation appears to be mediated by the corticosteroids, since injection of a combination of mineralocorticoid- and glucocorticoid receptor antagonists into the paraventricular thalamus before each of the repeated restraint bouts also blocked the acquisition of habituation
Amygdalar CRF synthesis and secretion is stimulated by glucocorticoid activity in amygdalar cells (3, 15–17) and CRF activity appears to be essential for the engagement of the chronic stress response network (4, 14)
reduction in plasma AVP is highly correlated with the reduction in corticosterone, and that injection of an AVP-neutralizing antiserum reduced corticosterone in water-deprived but not ad lib drinking rats
the splanchnic neural innervation of adrenocortical cells is of key importance in upregulation of adrenal sensitivity to ACTH diurnally (9) and to the response to water deprivation (19)
It seems likely that there is a similar mechanism in place under those conditions to rapidly relieve the organism of excessive corticosterone secretion once the stimulus is relieved. Limiting the increased corticosterone burden through a variety of mechanisms is physiologically important, and should serve to prolong life and more normal function under conditions of chronic challenge
van Dam D, van Nierop M, Viechtbauer W, Velthorst E, van Winkel R, GROUP investigators, Bruggeman R, Cahn W, de Haan L, Kahn R, Meijer C, Myin-Germeys I, van Os J, Wiersma D. 2014. “Childhood abuse and neglect in relation to the presence and persistence of psychotic and depressive symptomatology” Psychological Medicine 17: 1-15
http://www.ncbi.nlm.nih.gov/pubmed/25065372 in dropbox
patients ( n = 1119), their siblings ( n = 1057) and a control group ( n = 589)
more childhood trauma is associated with more severe psychopathology; stronger association for abuse and positive psychotic symptoms, no specific symptom association for neglect
D’Andrea W, Ford J, Stolbach B, Spinazzola J, van der Kolk B. 2012. “Understanding Interpersonal Trauma in Children: Why We Need a Developmentally Appropriate Trauma Diagnosis” American Journal of Orthopsychiatry 82(2): 187-200
http://www.ncbi.nlm.nih.gov/pubmed/22506521 in dropbox
Children exposed to interpersonal victimization often meet criteria for psychiatric disorders other than posttraumatic stress disorder (PTSD) … a wide range of symptoms is common in victimized children … in the current psychiatric nosology, multiple comorbid diagnoses are necessary—but not necessarily accurate—to describe many victimized children, potentially leading to both under-treatment and overtreatment
worldwide, approximately one third of children are estimated to experience physical abuse; approximately one in four girls and one in five boys experience sexual victimization
fewer than a quarter of children in treatment for trauma-related psychopathology with the National Child Traumatic Stress Network meet criteria for PTSD
Comorbidity is the rule: 40% of children with any trauma history have at least one other mood, anxiety, or disruptive behavior disorder diagnosis; this is exacerbated by exposure to
increasing numbers of types of traumatic stressors. … chronicity, physical violation, and betrayal of trust also determine risk and severity of posttraumatic symptoms and impairment, but a greater breadth of types of victimization is particularly influential in the development of multifaceted and severe symptoms
place at risk of chronic and severe coexisting problems with emotion regulation, impulse control, attention and cognition, dissociation, interpersonal relationships, and attributions
Although it may appear obvious that child maltreatment results in negative outcomes, current literature has remained fragmented; studies are conducted by islands of researchers who may not collaborate or integrate (e.g., experimental psychologists, epidemiologists, developmental psychologists, and clinicians)
the range of maltreatment, interpersonal violence, abuse, assault, and neglect experiences encountered by children and adolescents, including familial physical, sexual, emotional abuse and incest; community-, peer-, and school-based assault, molestation, and severe bullying; severe physical, medical, and emotional neglect; witnessing domestic violence; as well as the impact of serious and pervasive disruptions in caregiving as a consequence of severe caregiver mental illness, substance abuse, criminal involvement, or abrupt separation or traumatic loss
affective symptoms … exposed to interpersonal violence include lability, anhedonia, flat or numbed affect, explosive or sudden anger, and incongruous or inappropriate affect. Behavioral expressions of affect regulation may include withdrawal, self-injury, aggression, oppositional behavior, substance use, or other compulsive behavior. Behavioral dysregulation may represent affective overload as well as attempts to dispel, reduce, or recover from negative affect states
maltreated children are either hypersensitive or avoidant in response to negative emotional stimuli or are likely to interpret positive emotions as ambiguous
dissociation may take the form of inattention and impulsivity in traumatized children
domestic (esp interparental) violence increases risk and severity of internalizing, externalizing, relational, academic and vocational, and legal problems
maltreatment, sexual abuse, parental verbal abuse, and harsh corporal punishment have been found to be associated with numerous structural and functional alterations in the brain and neuroendocrine systems. Maltreated children have been found to have volumetric reductions in the corpus callosum left neocortex, hippocampus, and amygdala (Teicher et al., 2003). Young adult women who experienced sexual abuse, compared to matched controls, had reduced hippocampal volumes if the abuse occurred in early childhood or preadolescence, reduced corpus callosum volumes if the abuse occurred in middle childhood, and reduced prefrontal cortex volumes if the abuse occurred in adolescence (Andersen et al., 2008)
studies comparing women diagnosed with PTSD, depression, borderline personality disorder, and dissociative identity disorder who had childhood sexual abuse histories versus matched controls found evidence of reduced hippocampal (and in some cases, amygdalar) volumes (Bremner, Vithilingham, Vermetten, Southwick, et al., 2003; Schmahl, Vermetten, Elzinga, & Bremner, 2003; Vermetten, Sch-mahl, Lindner, Loewenstein, & Bremner, 2006; Vythilingam et al., 2002)
Women with childhood sexual abuse histories also have been shown to have reduced gray matter in the visual cortices (Tomoda, Navalta, Polcari, Sadato, & Teicher, 2009)
Harsh corporal punishment associated with reduced gray matter volumes in the medial and dorsolateral prefrontal cortices and anterior cingulate (Tomoda, Suzuki, et al., 2009)
Taylor et al. (2006) found that children who experienced harsh or cold parenting showed decreased amygdala activation during an emotion observation task and a strong relationship between amygdala activation and right ventrolateral prefrontal cortical areas during an emotion labeling task, which indicates poor inhibition of the amygdala
Young children who experienced abuse had lower cortisol than peers (King et al., 2001; Linares et al., 2008). women with childhood sexual abuse… similar neuroendocrine abnormalities (Bremner, Vermetten, & Kelley, 2007; Bremner, Vithilingam, Anderson, et al., 2003; Bremner, Vythilingam, Vermetten, et al., 2003).
The continued practice of applying multiple distinct comorbid diagnoses to traumatized children defies the cardinal rule of parsimony, obscures etiological clarity, and runs the danger of relegating trauma-informed treatment to only one disorder (PTSD) that is experienced by only a small fraction of traumatized children who are in psychiatric treatment
Dalton V, Long L, Weickert C, Zavitsanou K. 2011. “Paranoid schizophrenia is characterized by increased CB1 receptor binding in the dorsolateral prefrontal cortex” Neuropsychopharmacology 36(8): 1620-1630
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3138655/
paranoid SCZ having 22% higher levels of CB1R binding compared with the control group
Danese A, Moffitt T, Pariante C, Ambler A, Poulton R, Caspi A. 2008. “Elevated inflammation levels in depressed adults with a history of childhood maltreatment” Arch Gen Psychiatry 65(4): 409-415
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2923056/
Although depression was associated with high hsCRP (RR=1.45; 95%CI=1.06;1.99), this association was significantly attenuated and no longer significant when the effect of childhood maltreatment was taken into account. Individuals with current depression and childhood maltreatment history were more likely to show high hsCRP levels than controls (N=27; RR=2.07; 95%CI=1.23;3.47). In contrast, individuals with current depression only showed a non-significant elevation in risk (N=109; RR=1.40; 95%CI=0.97;2.01). Results generalized to the inflammation factor. The elevated inflammation levels in depressed+maltreated individuals were not explained by correlated risk factors, such as depression recurrence, low socioeconomic status in childhood or adulthood, poor health, or smoking
Danielson C, Amstadter A, Dangelmaier R, Resnick H, Saunders B, Kilpatrick D. 2009. “Trauma-related risk factors for substance abuse among male versus female young adults” Addict Behav 34(4): 395-399
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2704020/
1,753 young adults who participated in the 7−8 year follow-up telephone-based survey to the original National Survey of Adolescents. … 29.1% met criteria for substance abuse
exposure to specific traumatic events indeed increases risk for SUDs among young men. This is in contrast to much of the previous literature that has found that trauma exposure increases risk for SUDs among young women, but not young men. One possible explanation for this contrast is the inclusion of introductory narratives and behaviorally specific measurement of traumatic event history in the current study, which may have captured greater numbers of trauma-exposed young men than previous studies and provided a larger sample within which to examine the relation between such traumatic event history and SUDs
young adult women who have been sexually assaulted are at greater risk for alcohol abuse than men who have been sexually assaulted. Thus, gender may moderate the relation between alcohol abuse and SA
Dantzer R, O’Conner J, Freund G, Johnson R, Kelley K. 2008. “From inflammation to sickness and depression: when the immune system subjugates the brain” Nat Rev Neurosci 9(1): 46-56
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2919277/
In response to a peripheral infection, innate immune cells produce pro-inflammatory cytokines that act on the brain to cause sickness behaviour. When activation of the peripheral immune system continues unabated, such as during systemic infections, cancer or autoimmune diseases, the ensuing immune signalling to the brain can lead to an exacerbation of sickness and the development of symptoms of depression in vulnerable individuals
Darley C, Tinklenberg J, Roth W, Hollister L, Atkinson R. 1973. “Influence of marihuana on storage and retrieval processes in memory” Memory & Cognition 1(2): 196-200
http://link.springer.com/article/10.3758%2FBF03198094 add to dropbox
oral dose of marihuana extract calibrated to 20 mg of THC… One hour later. all Ss were administered delayed recall, recognition, and order tests on the first set of words, Presentation of another set of 10 lists followed. and there were immediate recall and delayed recall, recognition. and order t<:5!S on these words. Performance of drug and placebo Ss did not differ significantly for any of the first delayed tests. However. the performance of drug Ss was poorer than that of placebo Ss on immediate recall, delayed recall, and delayed recognition of the second set of lists. We concluded that retrieval of information relevant to the occurrence or nonoccurrence of an event was not affected by marihuana intoxication. Storage difficulties probably account for memory deficits due to the drug. and these difficulties appear to occur in the process of transferring information from short- term to long-term memory
The Ss were 48 adult males, all of whom were moderate(not more than once per week) social users of marihuana
Previous studies (Tinklenberg et al. 1970; Melges et aI, 1970: Abel. 1971a, b) have shown that marihuana intoxication causes decrements in the performance of certain memory tasks
Oral doses of the active drug were administered in the form of brownies containing NIMH marihuana extract calibrated to 20 mg of d9-tetrahydrocannibinol (THC)… required to fast for at least 8 h before the administration of the dose
it is important to ensure that the two groups did not differ by chance on the initial recall task
it is evident that not only is overall performance equal for the two groups, but probability of recall is virtually identical at each serial position
absence of differences between drug and placebo groups on delayed recall and recognition suggests that marihuana intoxication does not influence the retrieval of items stored in memory before drug intoxication. At least this is the case for the retrieval of information about whether or not a given item had been a member of a study list
Serial position curves for the immediate recall tests on the second set of lists are presented in the left panel of Fig. 2. The shapes of the curves are strikingly similar for the two groups, but performance for drug Ss is depressed for nearly all serial positions.
Drug and placebo Ss differ significantly in both delayed recall (placebo = .16, drug = .11, t(40) = 2.05, P < .05] and delayed recognition [placebo = .63, drug = .53, t(40) = 2.34, P < .05] . It is interesting to note that the groups differ even for terminal list positions in delayed recognition, even though the immediate and delayed recall curves converged for these items. On the second order-information test, the drug Ss again were less accurate in their estimates than were the placebo Ss. Meanabsolute deviations were 3.10 for the drug group and 2.97 for the placebo group. However,the difference between groups was not significant [t(40) = 1.08, p > .20] , and the fact that the mean values are based on different numbers of observations for the two groups (because of the different levels of delayed recognition performance) makes it difficult to interpret the result
Abel (1971a, b) performed a series of experiments using a procedure similar to that used here,and he found that marihuana intoxication had no effect uponrecall of previously stored information. However, performance on a delayed yes-no recognition test was adversely affected by the drug
if we are willing to conclude from the first part of the experiment that marihuana does not influence retrieval, then the differences observed in the second part of the experiment mustbe interpreted as being due to differential storage in the drug and placebo groups. Stated otherwise, marihuana has a decremental effect on the storage phase of memory, but has no effect on the retrieval stage
the apparent convergence in Fig. 2 of the immediate recall curves over terminal positions may indicate how storage isaffected bymarihuana. The recencyeffect for immediate recall can be explained as resulting from the near -perfect retrieval of terminal list items from short-term memory (Atkinson & Shiffrin, 1968, 1971). If it is the case that terminal items are in short-term memory at the time of test, then the fact that performance of drug and placebo groups for these items is similar may mean that drug Ss enterinformation into short-term memory as well as do placebo Ss
The results presented here indicate that during marihuana intoxication, Ss are not deficient in their ability to retrieve information which had been stored in memory before ingestion of the drug. In contrast, marihuana does affect storage processes so that information processed during intoxication is not remembered as well. It appears that marihuana does not influence the likelihood of entering information into short-term memory, but rather its transfer to a longer-term memory
Darley C, Tinklenberg J, Roth W, Vernon S, Kopell B. 1977. “Marijuana effets on long-term memory assessment and retrieval” Psychopharmacology 52, 239-241
http://link.springer.com/article/10.1007%2FBF00426706 add to dropbox
The ability of 16 college-educated male subjects to recall from long-term memory a series of common facts was tested during intoxication with marijuana extract calibrated to 0.3 mg/kg delta-9- tetrahydrocannabinol and during placebo conditions… cookies
Marijuana had no effect on recall or recognition performance… During both marijuana and placebo conditions, subjects could accurately predict their recognition memory performance
indicates that retrieval of non-experimentally-presented information from the long-term memory store is unaffected by marijuana. These data are consistent with previous findings that marijuana has no effect upon recall of word lists (Abel, 1971; Darley et al., 1973; Dornbush, 1974) or textual material (Miller et al., 1972) experimentally presented prior to drug intoxication
Our results suggest that subjects’ predictions of their own future memory task performance may be as accurate during marijuana intoxication as when they are unintoxicated
Davison S, Leese M, Taylor P. 2001. “Examination of the screening properties of the personality diagnostic questionnaire 4+ (PDQ-4+) in a prison population” J Pers Disord 15(2): 180-94
http://www.ncbi.nlm.nih.gov/pubmed/11345853/
There is a high prevalence of personality disorder in most prison populations. Many pass through the system undiagnosed
the prevalence of BPD was 45.7 percent by one measure, and, on the second, 47.4 percent
Day A, Metrik J, Spillane N, Kahler C. 2013. “Working memory and impulsivity predict marijuana-related problems among frequent users” Drug Alcohol Depend 131(0): 171-174
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3626751/
104 marijuana smokers marijuana use at least 2 days a week in the past month and at least weekly in the past 6 months, and self-reported ability to abstain from marijuana for 24 hours without withdrawal. Exclusion criteria were: intent to quit or receive treatment for cannabis abuse, use of other illicit drugs, pregnancy, nursing, past month affective or panic disorder, psychotic or suicidal state assessed by psychiatric interview, contraindicated medical issues by physical exam and BMI > 30, and smoking more than 20 tobacco cigarettes per day
Participants reported using marijuana a mean of 2.03 (SD = 1.22) times a day on 71.86% (SD = 22.02%) of the past 60 days (about 5 days per week). There were 44 tobacco cigarette smokers (42.3%), who smoked 4.08 cigarettes (SD = 3.77) per day. The 100 current drinkers drank 8.5 (SD=7.88) drinks per week consuming an average of 4.34 drinks (SD=2.44) on a drinking day
Although marijuana is the most commonly used illicit substance in the US, only a small portion of users go on to develop dependence, suggesting that there are substantial individual differences in vulnerability to marijuana-related problems among users. Deficits in working memory and high trait impulsivity are two factors that may place marijuana users at increased risk for experiencing related problems
Lower working memory, as measured by Trail Making Test B, but not short-term memory capacity, predicted more marijuana-related problems. Higher trait impulsivity scores were independently associated with greater number of problems
Participants most frequently reported that marijuana caused them “to procrastinate” (53% of the sample) and “to have a lower energy level” (42.6%). No participants reported that marijuana had caused them to lose a job, and 1% reported experiencing medical problems as a result of marijuana. The remaining 18 problems were reported by between 2 and 37% of participants
The current study did not examine motives or expectancies related to marijuana use, which have been shown to mediate the relation between use and subsequent problems (Mirin, 1971; Patrick et al., 2011;Simons et al., 1998)… negative expectancies about marijuana use have been shown to mediate the link between impulsivity and marijuana use (Vangsness et al., 2005), and as such, future studies might examine the role of motives expectancies in determining the link between impulsivity, executive functioning, use, and problems
Dean B, Sundram S, Bradbury R, Scarr E, Copolov D. 2001. “Studies on [3H]CP-55940 binding in the human central nervous system: regional specific changes in density of cannabinoid-1 receptors associated with schizophrenia and cannabis use” Neuroscience 103(1): 9-15
http://www.sciencedirect.com/science/article/pii/S0306452200005522
measured CB1R in dorsolateral prefrontal cortex (Brodmann’s area 9), caudate–putamen and areas of the temporal lobe from schizophrenic and control subjects; some used cannabis close to death. increase in the CB1R binding density in the dorsolateral prefrontal cortex from subjects with schizophrenia (mean±S.E.M.: 142±9.9 vs 119±6.6 fmol/mg estimated tissue equivalents; P<0.05) independent of recent cannabis ingestion. increase in density of CB1R in caudate–putamen from subjects who had recently ingested cannabis (151±9.0 vs 123±7.2 fmol/mg estimated tissue equivalents; P<0.05) that was independent of diagnoses.
De Bellis M, Keshavan M, Shifflett H, Iyengar S, Beers S, Hall J, Moritz G. 2002. “Brain structures in pediatric maltreatment-related posttraumatic stress disorder: a sociodemographically matched study” Biol Psychiatry 52(11): 1066-78
http://www.ncbi.nlm.nih.gov/pubmed/12460690
subjects with PTSD had smaller intracranial, cerebral, and prefrontal cortex, prefrontal cortical white matter, and right temporal lobe volumes and areas of the corpus callosum and its subregions (2, 4, 5, 6, and 7), and larger frontal lobe cerebrospinal fluid (CSF) volumes. The total midsagittal area of corpus callosum and middle and posterior regions remained smaller in subjects with PTSD, whereas right, left, and total lateral ventricles and frontal lobe CSF were proportionally larger
gender by group effect demonstrated greater lateral ventricular volume increases in maltreated males
larger prefrontal lobe cerebrospinal fluid volumes and smaller midsagittal area of the corpus callosum subregion seven (splenium)
De Bellis M, Kuchibhatla M. 2006. “Cerebellar volumes in pediatric maltreatment-related posttraumatic stress disorder.” Biol Psychiatry 60(7):697-703
http://www.ncbi.nlm.nih.gov/pubmed/16934769
Unadjusted means of the left, right, and total cerebellum were smaller in the PTSD group
De Bellis M, Spratt E, Hooper S. 2011. “Neurodevelopmental biology associated with childhood sexual abuse” J Child Sex Abuse 20(5): 548-587
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3769180/
Child maltreatment appears to be the single most preventable cause of mental illness and behavioral dysfunction in the US: disrupted development -> delays/deficits/failures of multisystem achievements in motor, emotional, behavioral, language, psychosocial, social, and cognitive skills
discussion of PTSD diagnosis, sub-clinical measures, differences in children
in clinically referred, PTSD incidence from sexual abuse range from 42% to 90% (Dubner & Motta, 1999; Lipschitz, Winegar, Hartnick, Foote, & Southwick, 1999; McLeer, Callaghan, Henry, & Wallen, 1994), from witnessing domestic violence from 50% to 100% (for domestic homicide; Pynoos & Nader, 1989), and from physical abuse to as high as 50% (Dubner & Motta, 1999; Green, 1985; Pynoos & Nader, 1989). few studies have assessed PTSD in non-clinically referred maltreated children. Famularo, Fenton and Kinscherff (1993) reported a 39% incidence rate of PTSD within eight weeks of abuse or neglect disclosure. About a third of the PTSD subjects re-examined from the original sample continued to meet PTSD criteria at two year follow-up (Famularo, Fenton, Augustyn, & Zuckerman, 1996). McLeer and colleagues (1998) reported prevalence rates of PTSD of 36.3%
Even if childhood trauma does not result in childhood PTSD, it increases the risk for adult PTSD (Bremner, Southwick, Johnson, Yehuda, & Charney, 1993; Widom, 1999) and health problems (Dube, Felitti, Dong, Giles, & Anda, 2003)
combat-related PTSD studies suggest that there is increased sensitivity of the locus ceruleus/SNS/catecholamine system, even in PTSD-
Elevated levels of corticotrophin releasing hormone (CRH) or factor (CRF); increased cortisol in children, decreased in adults.
sexually abused girls manifest a dysregulatory disorder of the HPA axis associated with hyporesponsiveness of the pituitary (ACTH) to exogenous CRH and normal overall cortisol secretion to CRH challenge
prepubertal abuse: larger pituitary. Pubertal/post-pubertal: smaller pituitary
Low serotonin function is associated with suicidal and aggressive behaviors In primate studies of chronic stress, serotonin levels decrease in the prefrontal cortex (Fontenot, Kaplan, Manuck, Arango, & Mann, 1995)
impaired immune, incl involution of the thymus and stress-induced lymphopenia
severe stress of sexual abuse may lead to suppression of the mechanisms (T suppressor cells) that actively suppress the autoantibody-producing lymphocytes (B lymphocytes) and may thus increase the incidence of positive antinuclear antibody titers in these sexually abused girls
medial prefrontal hyporesponsivity, amygdalar hyperresponsivity in PTSD.
monkeys were found to have a reduction in the midsagittal size of the corpus callosum, in parallel to a decrease in white (but not grey) matter volume in the prefrontal and parietal cortices
Teicher et al. (1997) early childhood trauma had reduction in the middle portion of the corpus callosum in children at psychiatric facilities with histories of trauma, including abuse or neglect, as compared to psychiatric controls (greater in males)
44 maltreated children and adolescents with PTSD and 61 age- and sex-matched controls, De Bellis (1999) decreased total midsagittal area of the corpus callosum and enlarged right, left, and total lateral ventricles in PTSD (males smaller CC, trend for smaller total brain) intracranial volume was decreased by 7% and total brain volume by 8% in PTSD subjects compared to controls; worse the earlier that abuse. PTSD symptoms correlated positively with ventricular volume
attenuation of frontal lobe asymmetry in children with maltreatment-related PTSD Carrion and colleagues (2001)
boys appear to be more sensitive to abuse in terms of brain development
maltreated children and adolescents with PTSD have lower NAA/creatine ratios suggests loss of neuronal integrity in the anterior cingulate region of the medial prefrontal cortex
maltreated subjects with PTSD had significantly larger, mainly right-sided, superior temporal gyrus grey matter volumes
intellectual ability (low IQ) may be due to child abuse. intellectual impairments leading to poor school performance (NationalResearchCouncil, 1993;Trickett & McBride-Chang, 1995), have been consistently reported in abused children not evaluated for PTSD (Augoustinos, 1987; Azar, Barnes, & Twentyman, 1988;Kolko, 1992; NationalResearchCouncil, 1993; Trickett & McBride-Chang, 1995). Negative correlations between Verbal IQ score and severity of abuse were observed (Carrey, Butter, Persinger, & Bialik, 1995). Perez & Widom (1994) reported lower IQ and reading ability in a large sample of adult survivors of child maltreatment
On average, children exposed to high levels of domestic violence had IQ scores eight points lower than children who were not exposed Koenen and colleagues (2003)
Decoster J, van Os J, Myin-Germeys I, De Hert M, van Winkel R. 2012. “Genetic variation underlying psychosis-inducing effects of cannabis: critical review and future directions” Current Pharmaceutical Design 18: 5015-5023
http://www.ncbi.nlm.nih.gov/pubmed/22716139
the relationship between cannabis use, cog-nition and psychosis is unclear
recent meta-analyses reported better cognition in cannabis-using patients [39, 40]. Several explanations for superior cognitive abilities in cannabis-using patients have been put forward, including possible neuroprotective effects of cannabis in individuals develop-ing psychotic symptoms [41] and a selective mechanism of causal contribution of cannabis, such that persons with less neurocognitive impairment make a transition to psychotic disorder
at the level of cognition, the evidence for differential sensitivity to cannabis associated with pre-existing liability is relatively inconsistent, for both the acute as well as the longer-term effects
greater reduction of grey matter density in cerebral regions known to have a high density of cannabinoid receptors (CB1R) in canna-bis-using patients [47-50], although this was not always found [51]. Ambiguous results were reported with regard to the effect of cannabis use on white matter integrity [47, 52]
COMT is an enzymatic inactiva-tor of dopamine and other mono-amines and is essential for dopa-mine signalling in the prefrontal cortex. The Val allele is associated with a 40% higher enzyme activity [58, 59]… interactions between cannabis and COMT showed greater variation, even with opposite directions in different analyses [66] [57-65]
It is disappointing that even the most studied gene-cannabis interaction (COMT Val158Met), has never been the subject of a true attempt at replication. It should be noted that the (negative) studies by Zammit and colleagues came closest to the original study design, in terms of phenotype under study [62] or in terms of the prospective design [66]
Defer N, Best-Belpomme M, Hanoune J. 2000. “Tissue specificity and physiological relevance of various isoforms of adenylyl cyclase” Renal Physiology 279 (3): F400-F416
http://ajprenal.physiology.org/content/279/3/F400.short
forty years after its discovery by Earl Sutherland, cAMP is still the archetypal “second messenger.” But the cAMP signaling pathway, once considered to be simple and straightforward, has become very complex indeed
mong the large diversity of the AC isoforms, some are widely expressed, such as AC2, AC4, and AC6, whereas others are more specifically expressed, for example AC1 in tissues of neural origin and AC5 in heart and striatum
AC1 is abundant in the dentate gyrus of the hippocampus and the cerebral cortex (189), the highest expression of AC3 is exhibited in the olfactory neuroepithelium (188), whereas the important area for AC8 expression is the hypothalamus (16,102), where it is the only Ca2+/calmodulin-stimulated isoform (110,187, 188)
All AC activities are inhibited by high, nonphysiological concentrations of Ca2+ in the submillimolar range, possibly by competition with magnesium
AC1 does function as a good coincidence detector, and this is well demonstrated in the pineal gland (170), where AC1 is activated by norepinephrine via both the β-AR (through Gsα) and the α1-AR (through Ca2+ release), to increase cAMP formation and ultimatelyN-acetyl transferase and melatonin synthesis
AC8 is also a major isoform in the brain although it has also been found in testis (36, 63) and lung (115). In the brain, it is mainly present throughout (16, 102, 115), especially in the cortex, cerebellum, brain stem, hypothalamus, hippocampus, and olfactory bulb. The specific localization in hypothalamic nuclei suggests a role in neuroendocrine function whereas its specific increase in some regions of the brain, and especially in the locus cœruleus during morphine administration and withdrawal, points to a role in drug dependence (87, 103)
It is interesting to note that another type of cyclase, AC9, is expressed to a high level in the brain
Acute administration of morphine or opioids causes a decrease of AC activity via the Gi pathway, and chronic administration leads to the classic states of tolerance and dependence (122)
After 6 days of treatment with Δ-9 tetrahydrocannabinol, followed by the administration of the antagonist, mice exhibit several somatic signs (wet-dog shakes, facial rubbing, ataxia, hunched posture, mastication) that could be interpreted as being part of a withdrawal syndrome. Interestingly, the same animals exhibit a 100% increase in the basal, forskolin-, and Ca2+/calmodulin-stimulated AC activity in the cerebellum (rich in CB1 receptors) but not in the cortex or the striatum (70).
In most cell culture systems, acute exposure to ethanol treatment has been found to potentiate the receptor-mediated cAMP synthesis. In contrast, chronic exposure often causes a decrease in cAMP production. Parsian et al. (127) have observed that basal and fluoride-stimulated platelet AC activity of alcoholic patients have lower value than in control subjects (127)
In human platelets, the major AC isoform expressed is AC7
Degenhardt L, Hall W, Lynskey M. 2002. “Alcohol, cannabis and tobacco use among Australians: a comparison of their associations with other drug use and use disorders, affective and anxiety disorders, and psychosis.” Addiction 96(11): 1603-1614
http://onlinelibrary.wiley.com/doi/10.1046/j.1360-0443.2001.961116037.x/abstract
nationally representative sample of 10 641 Australian adults
after controlling for demographics, neuroticism and other drug use, cannabis was not associated with anxiety or affective disorders. Alcohol dependence and tobacco use remained associated with both of these indicators of mental health
Degenhardt L, Hall W, Lynskey M. 2003. “Exploring the association between cannabis use and depression.” Addiction 98(11): 1493-1504
http://onlinelibrary.wiley.com/doi/10.1046/j.1360-0443.2003.00437.x/abstract
causation unclear… cannabis use makes, at most, a modest contribution to the population prevalence of depression.
little evidence of an increased risk of later cannabis use among people with depression and hence little support for the self-medication
Degenhardt L, Hall W, Lynskey M. 2003. “Testing hypotheses about the relationship between cannabis use and psychosis” Drug Alcohol Depend 71(1):37-48
http://www.ncbi.nlm.nih.gov/pubmed/12821204
There was a steep rise in the prevalence of cannabis use in Australia over the past 30 years and a corresponding decrease in the age of initiation of cannabis use. There was no evidence of a significant increase in the incidence of schizophrenia over the past 30 years. Data on trends the age of onset of schizophrenia did not show a clear pattern. Cannabis use among persons with schizophrenia has consistently been found to be more common than in the general population. Cannabis use does not appear to be causally related to the incidence of schizophrenia
Deiana S. 2013. “Medical use of cannabis. Cannabidiol: a new light for schizophrenia?” Drug Test Anal 5(1): 46-51
http://www.ncbi.nlm.nih.gov/pubmed/23109356?dopt=Abstract
Behavioural and neurochemical models suggest that CBD has a pharmacological profile similar to that of atypical anti-psychotic drugs and a clinical trial reported that this cannabinoid is a well-tolerated alternative treatment for schizophrenia.
DeLisi L, Bertisch H, Szulc K, Majcher M, Brown K, Bappal A, Ardekani B. 2006. “A preliminary DTI study showing no brain structural change associated with adolescent cannabis use.” Harm Reduct J 3: 17
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1524733/
cannabis use began prior to the age of 18 and consisted of use more than 21 times in any single year; no evidence of atrophy in any relevant regions
Demirakca T, Sartorius A, Ende G, Meyer N, Welzel H, Skopp G, Mann K, Hermann D. 2011. “Diminished gray matter in the hippocampus of cannabis users: Possible protective effects of cannabidiol” Drug and Alcohol Dependence 114 242-245
gray matter (GM) concentration and volume of the hippocampus were measured in 11 chronic recreational cannabis users and 13 healthy controls, and correlated with THC and CBD from hair analyses
Cannabis users showed lower GM volume located in a cluster of the right anterior hippocampus (P uncorr = 0.002; effect size Cohen’sd = 1.34). In a regression analysis an inverse correlation of the ratio THC/CBD with the volume of the right hippocampus (P uncorr p < 0.001, Cohen’s d = 3.43) was observed. Furthermore Cannabidiol correlated positively with GM concentration (unmodulated VBM data), but not with GM volume (modulated VBM) in the bilateral hippocampus
Cannabis was used for 5.4 years in an average daily dose of 0.27 g… Cannabis users were more likely to smoke tobacco cigarettes ( n = 6 versus n = 1) and consumed more alco-hol (21.4 g/day = about 1.5 drinks/day) than controls (4.2 g/day)
Group comparison of GM tissue composition or concentra-tion (unmodulated data) revealed no differences between healthy controls and cannabis users in whole brain or in hippocam-pal ROI
endocannabinoids promote oligoden-drocyte progenitor survival ( Molina-Holgado et al., 2002) and control axonal growth by inducing chemorepulsion or collapse of axonal growth cones ( Berghuis et al., 2007 ). Cannabionoid-1 (CB-1) receptor activation increases astroglial progenitor proliferation and differentiation in vitro, and neural progenitor proliferation and astrogliogenesis are decreased in adult CB1-deficient mice ( Aguado et al., 2006 )
Our results are limited by the small sample size and that volume differences did not survive a correction for the number of voxels in the hippocampus
Demuth D, Molleman A. 2006. “Cannabinoid signalling” Life Sciences 78: 549-563
http://www.ncbi.nlm.nih.gov/pubmed/16109430 add to dropbox
numerous studies have now shown that the signalling pathw ays utilised by cannabinoid receptors are a great deal more diverse than originally established
concentrations of cannabinoids that produced a profound sedation and inhibition of locomotor activity in mice did not reduce CB1 receptor binding by radioisotopes in the cerebellum and hippocampus (Gifford et al., 2002). This suggests that the occupancy of the CB1 receptor necessary for the behavioural effects of cannabinoids is very low
The CB1 cannabinoid receptor is a member of the rhodopsin subfamily of GPCRs. Cannabinoid agonists interact with the receptor within the pore formed within the trans membrane helical cluster (Mukhopadhyay et al., 2002). The three cytosolic loops and a putative fourth loop formed by palmitoylation at the juxtamembrane C-terminal region contribute to the activation of G proteins. The proxi mal CB1 receptor intracellular C- terminal domain (am ino aci ds 401 – 417) is critical f or G protein coupling and the distal C-terminal tail domain (amino acids 418 – 472) modulates the magni tude and kinetics of signal transduction ( Nie and Lewis, 2001)
Brain regions in which cannabinoids are effective inhibitors of adenylyl cyclase are those most densely populated with cannabinoid binding sites. Thus, cannabinoid-mediated inhibition of cAMP as been demonstrated in slices of rat hippocampus, striatum, cerebral cortex and cerebellum (Bidaut-Russell et al., 1990)
Nine distinct isozymes of adenylyl cyclase have been identified, which can be categorised into six distinct classes based on sequence and functional similarities (Patel et al, 2001): (a) AC type I (AC-I) is found mainly in brain, is stimulated by Ca2+/calmodulin and is inhibited by G beta gama subunits and by G i alpha; (b) AC-VIII is found only in brain and is only stimulated by Ca2+/calmodulin; (c) AC-II, AC-IV and AC-VII are activated by G beta gamma, providing that G s alpha is present; (d) AC-V and AC-VI are highly expressed in brain and heart, and they are inhibited by g i alpha and low levels of Ca2+; (2) AC-III is stimulated by a high concentration of Ca2+/calmodulin in the presence of G alpha s; (f) AC-IX is expressed at high levels in skeletal muscle and brain and, as yet, is found to be affected by G alpha s only
Monkey kidney COS-7 cells, expressing exogenous CB1 receptors, were transfected with each adenylyl cyclase isoform in turn and stimulated with the cannabinoid agonists HU-210 and WIN 55,212-2. AC-I, V, VI and VIII were shown to be inhibited by, whereas types II, IV and VII were stimulated by, CB1 receptor activation
the globus pallidus contains mRNA encoding for AC-II (Furuyama et al, 1993), an area of the brain where CB1 activation has been shown to induce an increase in cAMP accumulatin (Maneuf and Brotchie, 1997)
CB1 receptors have been shown to link positively to MAP kinase… human astrocytoma and CHO cells, expressing CB1 receptors, HU-210 and CP 55,940 activated a p42/p44 MAP kinase… in CHO cells was linked to the activation of a Na+/H+ exchanger (NHE-1), a transporter involved in multiple cellular functions such as intracellular pH regulation and control of cell volume (Bouaboula et al., 1999). In vivo, acute administration of d9-THC induces a progressive and transient activation of p42/p44 MAP kinase in rat dorsal striatum and nucleus accumbens Valijent et al., 2001), as well as in murine hippocampus (Derkinderen et al., 2003), striatum and cerebellum (Rubino et al., 2004). Blocked by CB1 antagonist… CP 55,940, WIN 55,212-2, anandamide and 2-AG have also been shown to stimulated p38 MAP kinase in rat and murine hippocampus
studies in primary rat astrocyte cultures showed that d9-THC and HU-210 increased glucose metabolism, phospholipid metabolism and glycogen synthesis through the activation of MAP kinase (Sanchez et al, 1998b)
Anandamide… acts via CB1 receptors to inhibit N-type VOCCs leading to a decrease in Ca2+ influx… inhibited Q-type Ca2+ currents in AtT-20 pituitary tumour cells expressing exogenous CB1 receptor… studies in rat cortical and cerebellar brain slices showed that anandamide inhibited P/Q-type Ca2+ fluxes
Activation of CB1 receptors by WIN 55,212-2 (nanomolar concentrations) inhibited only a fraction (17%) of the whole-cell CA2+ current, even though more than half of this current is carried by N- and P/Q-type Ca2+ channels (Shen and Thayer, 1998)… at micromolar concentrations the effects of WIN 55,212-2 are not mediated by CB1 receptors, which may suggest a direct effect of cannabinoids on Ca2+ channels. The inhibitory effect of cannabinoids on N-type Ca2+ channels in the hippocamus is in accordance with the observations of Wilson, 2001 who demonstrated that WIN 55,212-2 was unable to mimic depolarisation-induced suppression of inhibition in rat hippocampal slices pretreated with conotoxin
L-type Ca2+ channels can also be regulated via CB1 receptor stimulation
T-type Ca2+ channels transfected in HEK 293 and CHO cells and endogenously expressed in NG108-15 cells were inhibited by anandamide (Chemin et al, 2001) not mimicked by synthetic cannabinoids or blocked by antagonist
In cultured hippocampal neurons WIN 55,212-2 increased voltage-dependent A-type outward K+ currents (IsubA) (Deadwyler et al., 1995) and decreased voltage-independent D-type outward K+ currents (IsubD or delay current) (Mu et al., 1999) in a concentration-dependent, SR 141716A= and PTX sensitive manner. A cannabinoid-mediated reduction in cAMP/PKA was the mechanism shown to activate IsubA (Hampson 1995) and inhibit IsubD. It was proposed that phosphorylation of the K+ channel inactivated IsubA and therefore a decrease in PKA would act to reverse this process
in cerebellar granule and COS-7 cells anandamide inhibited the acid-sensitive background K+ channel TASK-1 (Maingret et al., 2001)
Nicholson et al (2003) demonstrated the ability of anandamide and WIN 55,212-2 to inhibit voltage dependent Na+ channels (activated by veratridine) in mouse synaptosomes. The cannabinoids also blocked teh veratridine-induced release of neurotransmitters from sypatosomes including GABA and glutamate. CB1 antagonst AM 251 did not attenuate sodium channel inhibition
In cultured human arterial endothelial cells anandamide evoked an increase in Ca2+i in an SR 141716A-sensitive manner… coupled to the production of NO… also in human umbilical endothelial cells only marginally blocked by SR, insensitive to PTX and blocked by caffeine suggesting a release of Ca2+ from caffeine-sensitive intracellular stores. Anandamide also significantly increased NO synthase activity… suggest that CB1 dependent and independent increases of Ca2+i and subsequent NO production may account for some of the vasodilator actions of anandamideit can be envisaged that Ca2+ release from intracellular stores in excitable cells can be inhibitory through activation of Ca2+ dependent K+ channels, leading to hyperpolarisation. This then would preevnt the activation of VDCC and large Ca2+ influx
Treatment of human prostate epithelial PC-3 cells with either THC or methanandamide activated the PI3K/PKB pathway, which in turn induced translocation of Raf-1 to the membrane and phosphorylation of p42/p44 MAP kinase… blocked by CB2 antagonist
cannabinoid-mediated reduction of MAP kinase may inhibit IL-2 production
on sensory neurons TRPV1 channel opening causes Ca2+ influx and neurotransmitter release… anandamide is thought to activate TRPV1 by binding to sites on the cytosolic side… Anandamide has also been shown to activate TRPV1 receptors in rat hippocampal slices
Nemeth (2003) investigated the effects of different concentrations of anandamide on neuropeptide release from sensory neurons of the rat trachea, which express both CB1 and TRPV1 receptors. Low concentrations of anandamide (10 microM) inhibited peptide release… high concentrations (50-100 microM) increased the release of peptides and this response was inhibited by capsazepine
cannabinoid mediated inhibition of 5-HT3 currents… cannabinoids do not interact directly with the agonist binding site of the 5-HT3 receptor, but may instead act allosterically at a 5-HT3 modulatory site
when the CB1 receptor component was blocked, anandamide but not d9-THC produced a stimulatory effect on NMDA-induced Ca2+ responses including rat cortical, cerebellar and hippocampal slices
cannabinoids may be able to modulate the activity of nAChRs in a cannabinoid receptor-independent manner
Denson TF, Earleywine M. 2006. “Decreased depression in marijuana users” Addictive Behaviour 31(4):738-42
http://www.ncbi.nlm.nih.gov/pubmed/15964704
” Despite comparable ranges of scores on all depression subscales, those who used once per week or less had less depressed mood, more positive affect, and fewer somatic complaints than non-users. Daily users reported less depressed mood and more positive affect than non-users. The three groups did not differ on interpersonal symptoms. Separate analyses for medical vs. recreational users demonstrated that medical users reported more depressed mood and more somatic complaints than recreational users, suggesting that medical conditions clearly contribute to depression scores and should be considered in studies of marijuana and depression. These data suggest that adults apparently do not increase their risk for depression by using marijuana.”
Denson T, Earleywine M. 2008. “Self-reported use of alcohol, marijuana, and hard drugs and aggression: a structural equation modeling analysis of an internet survey of long-term marijuana users” Journal of Aggression, Maltreatment and Trauma 16(2): 164-180
https://www.ncjrs.gov/App/publications/abstract.aspx?ID=245974
Although alcohol use and hard drug history were found to predict aggressive behavior, marijuana use did not
consistent with the majority of research that has shown no relationship between marijuana use and aggression
Deykin E, Buka S. 1997. “Prevalence and risk factors for posttraumatic stress disorder among chemically dependent adolescents” Am J Psychiatry 154(6): 752-7
http://www.ncbi.nlm.nih.gov/pubmed/9167501
297 adolescents aged 15-19 years who met the DSM-III-R criteria for dependence on alcohol or other drugs and who were receiving treatment
The lifetime prevalence of PTSD was 29.6% (24.3% for males and 45.3% for females), and the current prevalence was 19.2% (12.2% for males and 40.0% for females)
The lifetime prevalence of PTSD among these chemically dependent adolescents is five times that reported for a community sample of adolescents
De Petrocellis L, Ligresti A, Moriello A, Allara M, Bisogno T, Petrosino S, Stott C, Di Marzo V. 2011. “Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP chennels and endocannabinoid metabolic enzymes” British Journal of Pharmacology 163: 1479-1494
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3165957/
Diazgranados N, Ibrahim L, Brutsche N, Newberg A, Kronstein P, Khalife S, Kammerer W, Quezado Z, Luckenbaugh D. 2010. “A randomized add-on trial of an n-methyl-d-aspartate antagonist in treatment resistant bipolar depression” Arch Gen Psychiatry 67(8): 793-802
http://archpsyc.jamanetwork.com/article.aspx?articleid=210856
ketamine
Dickerson S, Gable S, Irwin M, Aziz N, Kemeny M. 2009. “Social-evaluative threat and proinflammatory cytokine regulation: an experimental laboratory investigation” Psychol Sci 20(10): 1237-1244
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2761517/
stimulated production of the proinflammatory cytokine tumor necrosis factor-α (TNF-α) increased from baseline to poststressor in the social-evaluative threat condition, but was unchanged in the non-SET condition. Further, the increases in TNF-α production correlated with participants’ cognitive appraisals of being evaluated. Additionally, the ability of glucocorticoids to shut down the inflammatory response was decreased in the SET condition. These findings underscore the importance of social evaluation as a threat capable of eliciting proinflammatory cytokine activity and altering its regulation.
Dill D, Chu J, Grob M, Eisen S. 1991. “The reliability of abuse history reports: a comparison of two inquiry formats.” Compr Psychiatry 32(2):166-9
http://www.ncbi.nlm.nih.gov/pubmed/2022116
nearly all reports of abuse histories obtained in the intake interview were consistent with later reports obtained in the survey. However, findings of no abuse history obtained in the intake format were frequently inconsistent with reports obtained in the survey, which were twice as frequent as intake reports.
Diseth T. 2006. “Dissociation following traumatic medical treatment procedures in childhood: a longitudinal follow-up” Dev Psychopathol 18(1): 233-51
http://www.ncbi.nlm.nih.gov/pubmed/16478561/
three groups of patients with congenital anomalies were examined longitudinally. At first admission, adolescents (ages 10-20, mean 15) with anorectal anomalies (n = 14), adolescents with Hirschsprung disease (n = 14), and hospitalized controls (n = 14)
Anal dilatation, an invasive medical treatment procedure performed daily by the parents the first 4 years, was correlated with the frequency and severity of persisting dissociative symptomatology. The procedure was the only significant predictor of A-DES and SDQ-20 scores, and one of two significant predictors of DES scores
impact of early traumatic exposure on child development in the absence of parental malevolence, and on later dissociative outcome in adolescence and adulthood
A trauma may be so intense and complex that the child’s mind is unable to process the events as a whole experience. The cognitive schemas are lacking, the level of affect is too high, and the traumatic experiences may become split off from conscious control. Dissociation (ie, a fragmentation of the experience) occurs. According to DSM-IV, pathological dissociation in children refers to a developmental “disruption in the usually integrated functions of consciousness, memory, identity, or perception of the environment” with psychological dissociative symptoms such as amnesia, depersonalization, derealization, fragmentation, absorption, and identity confusion. Children with dissociative failure may also have somatic symptoms that reflect somatoform dissociative phenomena such as sensory losses, loss of motor control, general paralysis, and alterations of vision, hearing, taste and smell (Nijenhuis, Spinhoven, Van Dyck, Van der Hart, & Vanderlinden, 1996)
strong association between a history of early, chronic abuse, and extensive dissociative experienecs in adults (Chu, 1999; Lange 1999), adolescents and children (Brunner, 2000; Kisiel, 2001). This research emphasizes the importance of age of onset, severity, chronicity; that is, the number of developmetal periods during which trauma was experienced, and the number of subtypes of the trauma experience
children may experience specific life-saving medical procedures and treatment, such as bone marrow transplanatation and chemotherapy, as traumatic
The early onset, daily, repetitive, long-lasting and invasive anal treatment procedure performed by the parents for years proved to be associated with the frequency and severity of mental health problems and dissociative symptoms. Duration… was the only significant predictor of both dissociative and mental health outcomes in adolescence, was still the only significant predictor of somatoform dissociative outcome in adults, and was one of two significant predictors of psychogical dissociative outcome in adults… similar to those indicative of a strong relationship between sexual and physical abuse and dissociation in children and adolescents (Kisiel, 2001; Nijenhuis, 1996)
chronicity Kendall-Tackett 1993
medical procedures are cited as traumatic events more often than any other disease-related traumatic event (Stuber, 2003)
at median of 7 months, children with HD were kept restrained and tied in bed for 5 days after an operation involving crushing clamps in the anorectum. In one-third of the HD children, a second procedure was performed at the average age of 1.5 years. The short-term, nonrepetitive restraints of teh child was reported by parents to be unpleasant for the child and parents alike, but no significant dissociative problems were reported as adolescents or adults. In children with ARA, anal dilatations were performed once or twice daily by the parents until the children reached an average age of 4 years. The parents reported taht the treatment had a traumatic effect on the children, causing power struggles and engendering parental guilt as a result of the discomfort and even pain inflicted on the child. Both the procedure itself and the children’s reactions to it may have affected the attitude of the parents and the parent-child relationship
the most severe traumatic experiences are those inflicted by the persons who were expected to be protectors… one aspect of the risk entailed by anal dilatation may be the absence of the sensitive and protective interaction between the parents and young child. The development of these executive functions requires maturation of the frontal lobes from the end of the first year (Glaser, 2000)
previous findings of low global self-esteem, school competence, and social acceptance in ARA adolescents compared with adolescents with other chronic physical disorders (Aasland, 1999). Also supported by the discovery of significant differences in A-DES, DES, SDQ-20 and IES scores in this sample between those who had anal dilatation up to 2.5 years of age and those who were treated with the procedure for more than 2.5 years
adult patients can better cope with their experiences of the traumatic treatment procedure, but for the adolescent, the experience is still so recent that coping is not yet possible
Treatment involving years of anal dilatation as a routine treatment procedure has no been brought to an end.
There is still much ignorance or, at best, discussions, about dissociative disorders in general. The criticism against today’s perception of dissociative disorders has focused on these disorders as iatrogenic and epidemic. … this study eliminates any doubt about whether repeated, long-term traumatization performed by caregivers has a long-lasting impact on the children’s development
Dixon L, Haas G, Welden P, Sweeney J, Frances A. 1990. “Acute effects of drug abuse in schizophrenia patients: clinical observations and patients’ self-reports” Schizophrenia Bulletin 16(1): 69-79
http://schizophreniabulletin.oxfordjournals.org/content/16/1/69.full.pdf
Though drug abuse may exacerbate psychotic symptoms, abused drugs may also lead to transient symptom reduction in subgroups of schizophrenic patients. Some patients report feeling less dysphoric, less anxious, and more energetic while intoxicated
If rational treatment strategies are to be designed, it is necessary to develop empirical and theoretical foundations to understand why schizophrenic patients abuse psychoactive drugs
Negrete et al. (1986) evaluated 137 schizophrenic outpatients and found that cannabis users had significantly more delusional and hallucinatory activity than nonusers. Heavy cannabis users showed the most symptomatology. Bernhardson and Gunne (1972) found that in 7 of the 14 patients whose psychosis predated cannabis use, cannabis abuse produced a further psychotic deterioration; cannabis did not appear to exert any effect on the remaining 7 patients.
clinical observations of drug-abusing schizophrenic inpatients are likely to overrepresent exacerbations and underrepresent benefits, since the study population is a relapsing one, and adverse effects are more likely to come to clinicians’ attention
Hekimian and Gershon (1968) reported that in their sample of eight cannabis users, of whom six had a predrug and postdrug diagnosis of schizophrenia, five reported a “favorable subjective response” to cannabis. Knudsen and Vilmar (1984) presented descriptions of 10 schizophrenic patients medicated with neuroleptics of whom 7, they contended, used cannabis to reverse neurolepric effects. For example, one patient reported that cannabis allowed her to “regain experiences blotted out by medication.” Knudsen and Vilmar described patients’ experiences with cannabis in detail; responses were characterized by an initial feeling of being “inspired,” “relaxed,” “energized,” or “active,” followed by an exacerbation of symptoms and feeling “bad,” “aggressive,” or “splitting of thoughts.”
Alcohol, cannabis, and cocaine were the drugs abused commonly enough for meaningful results to be obtained. The large majority of substance-abusing patients felt that all three drugs decreased depression (table 1). In contrast, reported effects on anxiety, energy, and psychotic symptoms differed for the three drugs. For example, drug-abusing patients reported that cannabis and alcohol decreased anxiety but that cocaine made them more anxious
dysphoria, anxiety, desire for socialization, and possibly the anergia associated with negative symptoms may impel schizophrenic patients to abuse drugs.
Is psychosis just an unwanted side effect of illicit drug use, or an intended effect?
a way of understanding the popularity of cannabis abuse is that cannabis, in addition to its reported acute antidepressant effects (common to cocaine and alcohol), was perceived to have desirable anxiolytic and activating effects
although evidence may suggest that schizophrenic patients self-medicate negative symptoms, negative symptoms presumably diminish the patients’ capacities to seek and obtain drugs. Thus, patients with the most severe negative symptoms would be unlikely to abuse drugs
dysphoria may be the final common pathway to drug abuse
Do Y, McKallip R, Nagarkatti M, Nagarkatti P. 2004. “Activation through cannabinoid receptors 1 and 2 on dendritic cells triggers NF-KB-dependent apoptosis: novel role for endogenous and exogenous cannabinoids in immunoregulation” Journal of Immunology 173(4): 2372-2382
http://www.jimmunol.org/content/173/4/2373.long
Addition of Δ9-tetrahydrocannabinol (THC), a major psychoactive component found in marijuana or anandamide, an endogenous cannabinoid, to DC cultures induced apoptosis in DCs. DCs expressed CB1 and CB2 receptors and the engagement of both receptors was necessary to trigger apoptosis. Treatment with THC induced caspase-2, -8, and -9 activation, cleavage of Bid, decreased mitochondrial membrane potential, and cytochrome c release, suggesting involvement of death-receptor and mitochondrial pathways. DCs from Bid-knockout mice were sensitive to THC-induced apoptosis thereby suggesting that Bid was dispensable. There was no induction of p44/p42 MAPK, p38 MAPK, or stress-activated protein/JNK pathway in THC-treated DCs. However, THC treatment induced phosphorylation of IκB-α, and enhanced the transcription of several apoptotic genes regulated by NF-κB. Moreover, inhibition of NF-κB was able to block THC-induced apoptosis in DCs. Lastly, in vivo treatment of mice with THC caused depletion of splenic DCs
Dorahy M, Corry M, Shannon M, Macsherry A, Hamilton G, McRobert G, Elder R, Hanna D. 2009. “Complex PTSD, interpersonal trauma and relational consequences: findings from a treatment-receiving Northern Irish sample” J Affect Disord 112(1-3): 71-80
http://www.ncbi.nlm.nih.gov/pubmed/18511130/
Complex PTSD is associated with PTSD but when present should be considered a superordinate diagnosis
Dorard G, Berthoz S, Phan O, Corcos M, Bungener C. 2008. “Affect dysregulation in cannabis abusers” Eur Child Adolsc Psychiatry 17: 274-282
http://www.ncbi.nlm.nih.gov/pubmed/18301941
32 cannabis abusers (CA) and 30 healthy controls completed a battery of self-reports measuring depression (BDI-13), anxiety (STAI-Y), alexi-thymia (TAS20; BVAQ-B), anhe-donia (PAS; SAS), and sensation seeking (SSS)… more than half of the CA reported at least one other non-drug or alcohol comorbid diagnosis. The most commmon were mood and anxiety disorders. CA subjects scored significantly higher on all affective dimensions except alexithymia total scores; however, they had greater scores for the two subscales measuring difficulties in identifying feelings
CA subjects were more likely to experience high levels of trait anxiety, physical anhedonia and sensation seeking than the controls.
puts emphasis on the importance of substance use prevention as early as middle school
various studies have demon-strated that daily or even regular cannabis use can massively affect cognitive abilities and motivation, and in turn ultimately impede educational achieve-ment, social adjustment, and emotional reactivity
recruited in the Centre Emergence — Espace Tolbiac addiction unit of the Institut Mutualiste Montsouris, Paris… the patients’ treatment started on request of the patients’ parents (56%), the patient him/herself (28%), the justice (12%) or a physician (4%)
Exclusion criteria were mental retardation, organic brain disease, chronic or severe somatic disorder with possible effect on psychological state, psychotic disorder, inability to read or fill out the questionnaires or refusal to participate
Among the CA group, 5 (15.6%) were in middle school (14-15 years old), 6 (18.8%) were in vocational school (14-18 years old) and 3 (9.4%) were in university (18 years old and more)
control group recruited among students attending a scholar drug use prevention program
8 CA subjects (26.7%) had comorbid current major depression, one had past major depressive episode, 6 had comorbid dysthymia over the previous 2 years, 3 had past hypomania, and 2 had past mania. Regarding anxiety disorders, 2 CA subjcts presented lifetime panic attack, 2 had current agoraphobia, 2 had current social phobia, and 6 had a generalized anxiety disorder over the previous 6 months. 18 met DSM-IV criteria for comorbid alcohol abuse, and 3 had alcohol dependence.
foremost drives for cannabis consumption were to achieve: a feeling of detachment (50%), relaxation and anxiety sedation (37.5%), and euphoria (12.5%)
Age of first use 13.3 +/- 1.3, max joints per day 15.3 +/- 8.5, current joints per day 4.9 +/- 3.1 days alcohol use per month 9.6 +/- 10.1, alcohol per day of use 4.1 +/- 2.5, cigarettes per day 12.3 +/- 7.6
Nine subjects had experienced ecstasy, 8 had used shrooms, 4 cocaine, 3 LSD, 3 sedatives, tranquilizers or pain relievers without medical prescription
25% of CA and 6.7% of controls had a severe depressive symptomatology, 37.5% of CA and 16.7% of controls had a moderate depressive symptomatology, 28.1% of CA and 23.3% of controls had a mild depressive symptomatology, and 9.$% of CA and 53.3% of controls had no depressive symptomatology… 40.6% of CA and 24.1% of controls categorized as alexithymic, 37.5% of CA and 48.3% of controls were intermediate, and 21.9% of CA and 27.6% of controls were non-alexithymic
Although the SSS total score and subscores were not correlated with the cannabis use modalities, we observed a positive correlation between the SSS total score and the number of days of alcohol use on the previous month and the age of first tobacco use
an 85% psychiatric comorbidity rate has been assessed through the cannabis abusers’ sample, when excluding psychotic patients.
Although causality linkage could not be established in this cross sectional study, rates and levels of depressive and anxiety symptomatology of the CA subjects suggest a specific association between cannabis dependence and mood disorders and symptoms
the fact that CA subjects’ motivation for cannabis use was mainly detachment and anxiety sedation, togetehr with the finding of a high prevalence of lonely cannabis and alchol use, might give some additional support to the self-medication model… higher physical anhedonia and sensation seeking scores of the CA
the sample’s representativeness is questionable
Dube SR, Felitti VJ, Dong M, Chapman DP, Giles WH, Anda RF. 2003. “Childhood abuse, neglect, and household dysfunction and the risk of illicit drug use: the adverse childhood experiences study.” Pediatrics 111(3):564-72
http://www.ncbi.nlm.nih.gov/pubmed/12612237
“the effects of adverse childhood experiences transcend secular changes such as increased availability of drugs, social attitudes toward drugs, and recent massive expenditures and public information campaigns to prevent drug use. … [adverse childhood experiences] seem to account for one half to two third of serious problems with drug use”
The attributable risk fractions as a result of ACEs for each of these illicit drug use problems were 56%, 64%, and 67%, respectively.
Durkee C, Sarlls J, Hommer D, Momenan R. 2013. “White Matter Microstructure Alterations: A Study of Alcoholics with and without Post-Traumatic Stress Disorder.” PLOS ONE doi: 10.1371/journal.pone.0080952
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0080952 in dropbox
Harris et al. [19], lower FA in alcoholics in the right superior longitudinal fascicles II and III, orbitofrontal cortex white matter, and cingulum bundle associated with lower working memory. Pfefferbaum et al. [20] showed that the greatest white matter abnormalities in alcoholics compared to healthy controls were observed in the frontal forceps, internal and external capsules, fornix, and longitudinal fasciculus, and that these were correlated with poorer performance on psychomotor speed tests. Other white matter regions typically implicated in alcoholism are the frontal and limbic pathways, centrum semiovale, and corpus callosum [11,20–24], with the genu of the corpus callosum and other more anterior white matter structures often found to be more compromised than posterior structures [20,25]
Durrant J, Ensom R. 2012. “Physical punishment of children: lessons from 20 years of research” CMAJ 184(12): 1373-1377
http://www.cmaj.ca/content/184/12/1373 in dropbox
Shift in public policy and perspective since 1990; physical punishment now widely recognised as unacceptable
Physical punishment increases aggression; reduction in harsh discipline was followed by reduced aggression
a range of mental health problems in children, youth and adults, including depression, unhappiness, anxiety, feelings of hopelessness, use of drugs and alcohol, and general psychological maladjustment
These relationships may be mediated by disruptions in parent–child attachment resulting from pain inflicted by a caregiver,30,31 by increased levels of cortisol32 or by chemical disruption of the brain’s mechanism for regulating stress.33 Researchers are also finding that physical punishment is linked to slower cognitive development and adversely affects academic achievement.
Driessen M, Herrmann J, Stahl K, Zwaan M, Meier S, Hill A, Osterheider M, Petersen D. 2000. “Magnetic resonance imaging volumes of the hippocampus and the amygdala in women with borderline personality disorder and early traumatization” Arch Gen Psychiatry 57(12):1115-22
http://archpsyc.jamanetwork.com/article.aspx?articleid=481684
patients with BPD had nearly 16% smaller volumes of the hippocampus (P<.001) and 8% smaller volumes of the amygdala (P<.05) than the healthy controls. The results for both hemispheres were nearly identical
hippocampal volume negatively correlated with extent and duration of self-reported early traumatization only when BPD and control subjects were considered together. Levels of neuropsychological functioning were associated with the severity of depression but not with the volumes of the hippocampus.
Edwards C, Skosnik P, Steinmetz A, Vollmer J, O’Donnell B, Hetrick W. 2008. “assessment of forebrain-dependent trace eyeblink conditioning in chronic cannabis users” Neuroscience Letters 439: 264-268
http://www.ncbi.nlm.nih.gov/pubmed/18534754 add to dropbox
While CB1 knockout mice exhibit striking impairments on a cerebellar-dependent task called delay eyeblink conditioning (dEBC), these animals demonstrate intact forebrain-dependent trace EBC (tEBC)… 13 heavy cannabis users and 13 cananbis naive controls completed a tEBC procedure… exhibited similar rates of conditioned responding compared to controls in the acquisition and extinction phase. Consistent with reports of overt attentional abnormalities, the cannabis group exhibited decreased N100 ERP amplitudes to teh tone CS that were unrelated to mean levels of conditioning across blocks during the acquisition phase
Skosnik demonstrated that chronic cannabis users are similarly impaired on the dEBC procedure (as knockout mice, as shown by Kishimoto and Kano)
All inclusion criteria, clinical interviews, drug use assessment, experimental stimuli, procedures and data processing methods are identical to those employed by Skosnik, with the exception of the 500 ms trace period
minimum 1 joint per week, 24hr abstinence, no DSM-IV axis I or II diagnosis except cannabis abuse or dependence
mean 6.7 joints per week, 44 per month, 228 ppast six months, age of onset 15.46, years of use 3.2, alcoholic drinks per week 5, cigarettes per day 1.6 (4.25) (no alcohol or cigarette use by controls)
covarying for alcohol consumption on % CRs did not alter the sig-nificant main effect of group, and a Pearson correlation between the reported average number of cigarettes smoked per day within the cannabis and the mean % CRs yielded a non-significant relationship, r(13) = .11, p =.73
The total awareness score did not differ between the groups… Nor were the scores significantly associated with change in percent CRs across the acquisition phase… group rates of extinction did not differ
N100 amplitudes were significantly smaller in the cannabis group (mean = −5.23, S.D. = 2.68) compared to the control group (mean = −8.27, S.D. = 3.81) ( F(1, 24) = 5.25, p = .031, d = .92; seeFig. 1B). N100 latencies, however, did not differ between the cannabis group (mean = 128.22, S.D. = 16.99) and the control group (mean = 129.39, S.D. = 12.92) ( F(1, 24) = .04, p = .85, d = .08)
the rate of cigarette smoking, within the cannabis group, was significantly associated with N100 latency ( r(25) = −.71, p = .01)
tEBC appears to recruit a broader forebrain network including the anterior cingulate, dorsolateral prefrontal cortex, hippocampus, caudate, and sensory cortex in order to properly encode the trace interval
the present N100 finding differs from the results of a recent study assessing dEBC in cannabis users in which no differences in the N100 to the CS were identified
Ehlers C, Slutske W, Lind P, Wilhelmsen K. 2007. “Association between single nucleotide polymorphisms in the cannabinoid receptor gene (CNR1) and impulsivity in southwest California Indians” Twin Res Hum Genet 10(6): 805-11
http://www.ncbi.nlm.nih.gov/pubmed/18179391
Ehrenreich H, Rinn T, Kunert H, Moeller M, Poser W, Schilling L, Gigerenzer G, Hoehe M. 1999. “Specific attentional dysfunction in adults following early start of cannabis use.” Psychopharmacology (Berl) 142(3):295-301
http://www.ncbi.nlm.nih.gov/pubmed/10208322
Of the potential predictors of test performance within the user group, including present age, age of onset of cannabis use, degree of acute intoxication (THC+THCOH plasma levels), and cumulative toxicity (estimated total life dose), an early age of onset turned out to be the only predictor, predicting impaired reaction times exclusively in visual scanning. Early-onset users (onset before age 16; n = 48) showed a significant impairment in reaction times in this function, whereas late-onset users (onset after age 16; n = 51) did not differ from controls (n = 49).
Ehrhart J, Obregon D, Mori T, Hou H, Sun N, Bai Y, Klein T, Fernandez F, Tan J, Shytle R. 2005. “Stimulation of cannabinoid receptor 2 (CB2) suppresses microglial activation” J Neuroinflammation 2: 29
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1352348/
Activated microglial cells have been implicated in a number of neurodegenerative disorders, including Alzheimer’s disease (AD), multiple sclerosis (MS), and HIV dementia. It is well known that inflammatory mediators such as nitric oxide (NO), cytokines, and chemokines play an important role in microglial cell-associated neuron cell damage. Our previous studies have shown that CD40 signaling is involved in pathological activation of microglial cells. Many data reveal that cannabinoids mediate suppression of inflammation in vitro and in vivo through stimulation of cannabinoid receptor 2 (CB2)
selective stimulation of cannabinoid receptor CB2 by JWH-015 suppressed IFN-γ-induced CD40 expression. In addition, this CB2 agonist markedly inhibited IFN-γ-induced phosphorylation of JAK/STAT1. Further, this stimulation was also able to suppress microglial TNF-α and nitric oxide production induced either by IFN-γ or Aβ peptide challenge in the presence of CD40 ligation. Finally, we showed that CB2 activation by JWH-015 markedly attenuated CD40-mediated inhibition of microglial phagocytosis of Aβ1–42 peptide. Taken together, these results provide mechanistic insight into beneficial effects provided by cannabinoid receptor CB2 modulation in neurodegenerative diseases
Ligation of CD40 on microglial cells leads to the production of TNF-α and other unidentified neurotoxins [11–13]
In the normal brain, microglial cells display a quiescent phenotype, including low CD40 expression [14]. However, upon insult to the brain, microglial cells become highly activated, altering their phagocytic and antigen-presentation functions [15] as well as the production of cytokines [13]. Mounting evidence implicates microglial CD40 as contributing to the initiation and/or progression of several neurodegenerative diseases [15]
the treatment of cultured microglial cells with THC, CP55940 and JWH-015 significantly inhibited CD40 expression induced by IFN-γ
stimulation of CB2 decreases CD40 expression on primary cultured microglial cells
CB2 protein is detected in primary cultured microglial cells, and is also markedly increased following the challenge with IFN-γ
either IFN-γ /CD40L or Aβ1–42/CD40L increased the secretion of the pro-inflammatory molecules TNF-α and NO, as indicated in Figure and . However, when CB2 is stimulated by the presence of JWH-015, these pro-inflammatory molecules were significantly reduced
increased numbers of CD40-positive microglial cells as well as increased CD40L expression on astrocytes in AD [53,54]
Aβ peptide can synergize with the IFN-γ signaling pathway to induce microglial CD40 expression and subsequent neurotoxicity [28]
CB2 agonist JWH015 inhibits IFN-γ-induced microglial CD40 expression by opposing JAK/STAT1 pathway activation
Eldreth D, Matochik J, Cadet J, Bolla K. 2004. “Abnormal brain activity in prefrontal brain regions in abstinent marijuana users” Neuroimage 23(3): 914-20
http://www.ncbi.nlm.nih.gov/pubmed/15528091 in dropbox
25-day abstinent, heavy marijuana users (n = 11), and a matched comparison group (n = 11); no deficits in performance on modified Stroop
hypoactivity in the left perigenual anterior cingulate cortex (ACC) and the left lateral prefrontal cortex (LPFC) and hyperactivity in the hippocampus bilaterally
El-Gabalawy H, Guenther L, Bernstein C. 2010. “Epidemiology of immune-mediated inflammatory diseases: incidence, prevalence, natural history and comorbidities” J Rheumatol Suppl 85: 2-10
http://www.ncbi.nlm.nih.gov/pubmed/20436161
Immune-mediated inflammatory diseases (IMID) present a group of common and highly disabling chronic conditions that share inflammatory pathways. Several incidence and prevalence studies of IMID during the past decades have reported a considerable variation of the disease occurrence among different populations. Overall, the estimated prevalence of IMID in Western society is 5%-7%
Elsey J, Coates A, Lacadie C, McCrory E, Sinha R, Mayes L, Potenza M. 2015. “Childhood trauma and neural responses to personalized stress, favorite-food and neutral-relaxing cues in adolescents” Neuropsychopharmacology doi:10.1038/npp.2015.6
http://www.nature.com/npp/journal/vaop/ncurrent/full/npp20156a.html
Relative to the low-trauma-exposed group, high-trauma-exposed adolescents displayed an increased activation of insula, anterior cingulate, and prefrontal cortex in response to stress cues. Activation in subcortical structures, including the hippocampus, was inversely correlated with subjective anxiety in the high- but not the low-trauma-exposed group. The high-trauma-exposed group displayed hypoactivity of cerebellar regions in response to neutral/relaxing cues. No group differences were observed in response to favorite-food cues
Elvevåg B, Goldberg T. 2000. “Cognitive impairment in schizophrenia is the core of the disorder” Crit Rev Neurobiol 14(1): 1-21
http://www.ncbi.nlm.nih.gov/pubmed/11253953/
we argue that various cognitive deficits are enduring features of the schizophrenia illness, that they are not state-related and are not specific to subtypes of the illness, and, more specifically, that working memory and attention are characteristically impaired in patients with schizophrenia, irrespective of their level of intelligence. Last, we conclude that problems in these cognitive domains are at the very core of the dysfunction in this disease
Elzinga B, Bremner J. 2001. “Are the neural substrates of memory the final common pathway in posttraumatic stress disorder (PTSD)?” Journal of Affective Disorders 70: 1-17
http://www.ncbi.nlm.nih.gov/pubmed/12113915 in dropbox
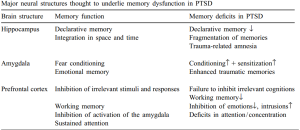
the noradrenergic stress-system is involved in enhanced encoding of emotional memories, sensitization, and fear conditioning, by way of its effects on the amygdala
two types of memory disturbances have been identified in traumatized individuals; intrusive memories and impoverished memory functioning
patients with PTSD develop serious distortions in the way they process information
trauma provides highly salient information that does not fit into preexisting schema’s or models of the world (Foa et al., 1989; Brewin et al., 1996)
PTSD is not only characterized by intrusive memories, but is also associated with general deficits in declarative memory, fragmentation of memories (both autobiographical and trauma-related), and trauma-related amnesia. Even though intrusions and amnesia may appear to be opposite phenomena these may instead be interrelated processes in that the occupancy with intrusive memories may interfere with, and thus reduce, memory processing of other material
Vietnam veterans with combat-related PTSD scored significantly lower (almost 50%) on the Weschsler Memory Scale (WMS) (Bremner et al., 1993a)
More than 30% of adults who had been sexually abused as children either failed to report or were amnesic for the event many years later (Williams, 1994)
subjects with high scores on dissociative and imaginative capacities appeared to be more prone to create false memories
Women with abuse-related PTSD had a higher frequency of false recognitions of critical lures then women with abuse histories without PTSD, non-abused non-PTSD women, or men without abuse or PTSD (Bremner, 2000)
Thus, PTSD-patients may be more prone for the creation of false memories, not so much because they are more suggestible, but because of their general memory deficits, leaving more room for elaboration
Dissociation is defined as a disruption in the usually integrated functions of consciousness, memory, identity or perception of the environment… dissociative reactions, such as rerealisation or depersonalisation, may impede memory encoding, so that traumatic memories can not be stored properly
amnesia and related dissociative symptoms at the time of trauma are associated with a long-term increase in psychopathology (Bremner et al., 1992; Marmar et al., 1994; Shalev et al., 1996; Bremner and Brett, 1997a)
Adrenaline has a short-term effect to strengthen memory traces, while cortisol has a more long-term effect to inhibit the laying down of memory traces
The noradrenergic system is responsible for rapid responses to stress, including the fight or flight response. Chronic stress is associated with increased firing of noradrenaline neurons in the brainstem and potentiated release of noradrenaline in the brain with subsequent stressors. (Bremner et al., 1996b) The noradrenergic system strengthens the formation of memory traces associated with emotional events.
memory enhancing effects of emotionally arousing events are mediated by the activation of b-adrenergic activity within the amygdala complex (Cahill and McGaugh, 1998)
when retrieval of a traumatic event is accompanied by adrenaline release, this may further strengthen the traumatic memory trace. A positive feedback loop may then result in deeply engraved memories, which are clinically expressed as intrusive recollections and flashbacks that are difficult to erase
lactate or yohimbine, substances that activate the noradrenergic system, induced intrusive memories and flashbacks in combat veterans with PTSD, besides other PTSD-symptoms including emotional numbing, and panic attacks (Southwick et al., 1993).
Noradrenaline appears to have opposite actions at alpha-1 and alpha-2 receptors; noradrenaline can impair PFC function through its actions at post-synaptic alpha-1 adrenergic receptors, whereas actions at alpha-2 receptors may improve performance. Since noradrenaline has higher affinity for alpha-2 than for alpha-1 receptors, low levels of noradrenaline release may engage alpha-2 receptors, whereas higher concentrations may engage alpha-1 receptors, resulting in impaired PFC function, including impaired working memory and response inhibition (Arnsten, 1998)
medial PFC dysfunction may be partially responsible for the failure to extinguish fear
Because of its glucocorticoid (GC) receptor sites, the hippocampus is the primary site of feedback for GC regulation keeping cortisol levels within physiological range. This makes the hippocampus particularly sensitive to stress
hipocampal atrophy may provide an explanation for the general memory deficits of PTSD patients
a differential effect was found between procedural and declarative memory: cortisol did impair declarative, but not procedural, memory (Kirschbaum et al., 1996; Lupien et al., 1997; Newcomer et al., 1999)
memory dysfunction as a core element of PTSD. While witnessing a traumatic event two stress reactions may occur. As a first rapid reaction, (nor)adrenaline is secreted. This may strengthen emotional memory traces, and enhance fear conditioning. Since the retrieval of traumatic memories is often accompanied by enhanced levels of adrenaline, this may further engrave the traumatic memory each time that it is retrieved. … Dysfunction of the medial PFC, a structure that normally inhibits the activation of the amygdala, may further enhance the effects of amygdala function, thereby increasing the emotional valence and the frequency of intrusive memories.
Cortisol may result in acute (but reversible) hippocampal dysfunction, an thus in verbal, declarative memory deficits, such as trauma-related amnesia, while leaving non-hippocampus mediated memory processes unaffected. Chronic cortisol secretion may eventually result in a permanent loss or atrophy of hippocampal cells. … dysfunction or atrophy of the hippocampus may also underlie distortions and fragmentations of trauma memories
Lack of self-esteem has been found to correlated with cortisol response in a stress task (Pruessner, 1999)
In animal models, adrenal activity has been related more to “defense” reactions, whereas cortisol is more associated with “defeat” (Henry and Stephens, 1977)
it must be stressed that… reconstructed memories are dependent on the emotions and needs of the context in which they are retrieved (Schacter, 1995)
Emeny R, Lacruz M, Baumert J, Zierer A, von Eisenhart Rothe A, Autenrieth C, Herder C, Koenig W, Thorand B, Ladwig K. 2012. “Job strain associated CRP is mediated by leisure time physical activity: results from the MONICA/KORA study” Brain Behav Immun 26(7): 1077-84
http://www.ncbi.nlm.nih.gov/pubmed/22813435
High job strain was reported by half (n = 482, 50.7%) of the study participants. While workers with high job strain were more likely to have adverse workplace conditions (competition with coworkers, job dissatisfaction and insecurity), sleeping problems, depressive symptoms, a Type A personality, and be physically inactive, no differences in cardiometabolic risk factors were detected. A strong and robust association between job strain and C-reactive protein (CRP) was observed in age and sex adjusted models, as well as models adjusted for classic coronary heart disease risk factors (β = 0.39, p = 0.006 and β = 0.27, p = 0.03, respectively). Adjustment for physical activity abrogated this effect (β = 0.23, p = 0.07), and a mediating effect of physical activity on stress-associated inflammation was demonstrated (p = 0.04)
Endo T, Sugiyama T, Someya T. 2006. “Attention-deficit/hyperactivity disorder and dissociative disorder among abused children” Psychiatry Clin Neurosci 60(4): 434-8
http://www.ncbi.nlm.nih.gov/pubmed/16884444
39 abused child inpatients in Aichi Japan. 59% dissociative disorder, 18% diagnosed ADHD, 71% of whom dissociative disorder. 67% met ADHD criteria, but only 27% prior to child abuse. Those with dissociative disorder increased apparent ADHD.
England T, Hind W, Rasid N, O’Sullivan S. 2015. “Cannabinoids in experimental stroke: a systematic review and meta-analysis” J Cereb Blood Flow Metab 35(3): 348-58
http://www.ncbi.nlm.nih.gov/pubmed/25492113/
In all, 144 experiments (34 publications) assessed CBs on infarct volume in 1,473 animals. Cannabinoids reduced infarct volume in transient (SMD -1.41 (95% CI -1.71), -1.11) P<0.00001) and permanent (-1.67 (-2.08, -1.27), P<0.00001) ischemia and in all subclasses: endocannabinoids (-1.72 (-2.62, -0.82), P=0.0002), CB1/CB2 ligands (-1.75 (-2.19, -1.31), P<0.00001), CB2 ligands (-1.65 (-2.09, -1.22), P<0.00001), cannabidiol (-1.20 (-1.63, -0.77), P<0.00001), Δ(9)-tetrahydrocannabinol (-1.43 (-2.01, -0.86), P<0.00001), and HU-211 (-2.90 (-4.24, -1.56), P<0.0001). Early and late neuroscores significantly improved with CB use (-1.27 (-1.58, -0.95), P<0.00001; -1.63 (-2.64, -0.62), P<0.002 respectively) and there was no effect on survival. Statistical heterogeneity and publication bias was present, median study quality was 4 (range 1 to 6/8). Overall, CBs significantly reduced infarct volume and improve functional outcome in experimental stroke
Englund A, Morrison P, Nottage J, Hague D, Kane F, Bonaccorso S, Stone J, Reichenberg A, Brenneisen R, Holt D, Feilding A, Walker L, Murray R, Kapur S. 2012. “Cannabidiol inhibits THC-elicited paranoid symptoms and hippocampal-dependent memory impairment” J Psychopharmacol 27: 19
http://www.ncbi.nlm.nih.gov/pubmed/23042808 add to dropbox
There is a wide inter-individual variation in PANSS positive scores following THC and, as a group, positive symptoms are modest compared with acute schizophrenia. In ear -lier studies approximately 35–50% of healthy participants showed changes of ≥3–4 points (D’Souza et al. 2004; Morrison et al., 2009)
In the placebo group, PANSS posi-tive scores, (mean±sd) increased by 2.4 (±3.1) points following THC, compared with 1.2 (±1.8) in the CBD group, a non-signifi -cant difference ( t =1.5, p =0.15) Clinically significant positive symptoms following THC, defined as an increase in PANSS positive scores of ≥ 3 points, were more common in the group pre-treated with placebo (11 of 26 cases) compared with the group pre-treated with CBD (3 of 22 cases), ( χ 2=4.74, p<0.05) (Table 3)
Energetic arousal decreased in the CBD group following the administration of CBD (p<0.01), whereas subsequent decreases following THC were not significant ( p=0.13)
Following THC, immediate recall was 2.9 (±5.3) and 3.6 (±4.5) items fewer in the CBD and placebo groups, respectively, compared with baseline
pre-treatment with CBD inhibited THC-induced paranoia and inhibited the detrimental effects of THC on episodic memory. In addition, CBD decreased the pro -portion of participants who experienced clinically significant acute THC psychosis… CBD (in so far as it was administered here) does not completely abolish THC-induced positive psychotic symptoms.
Here there were marked performance deficits post-THC in three tests which require pre-frontal resources: immediate recall, digit-span forward and digit-span back. CBD did not appear to attenuate THC-induced deficits in any of the three tasks. This contrasted with the protective effect of CBD on delayed recall and paranoid symp -toms. It is also notable that THC-induced impairment in delayed recall and THC-induced paranoia were correlated, and it is feasible that both measures load onto a common factor
Etkin A, Wager T. 2007. “Functional neuroimaging of anxiety: a meta-analysis of emotional processing in PTSD, Social Anxiety Disorder, and Specific Phobia” Am J Psychiatry 164(10): 1476-1488
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3318959/
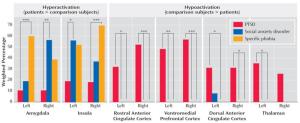
Patients with any of the three disorders consistently showed greater activity than matched comparison subjects in the amygdala and insula… similar pattern was observed during fear conditioning in healthy subjects… only patients with PTSD showed hypoactivation in the dorsal and rostral anterior cingulate cortices and the ventromedial prefrontal cortex—structures linked to the experience and regulation of emotion
Despite some shared key features, anxiety disorders also differ in a number of fundamental ways. Symptoms of hypervigilance and hyperarousal, dissociation, emotional numbing, and reexperiencing phenomena (nightmares and flashbacks) are particularly characteristic of PTSD (1). These symptoms are not observed during normal fear conditioning, suggesting that PTSD involves either different or more profound emotional dysregulation than other anxiety disorders
Hypoactivations (comparison subjects > patients) were seen only in PTSD, specifically in the inferior occipital gyrus, ventromedial prefrontal cortex, rostral anterior cingulate cortex, parahippocampal gyrus, lingual gyrus, dorsal amygdala and anterior hippocampus, orbitofrontal cortex, putamen, middle occipital gyrus, dorsomedial prefrontal cortex, dorsal anterior cingulate cortex, and mid-cingulate
for the thalamus, hypoactivity was observed more frequently than hyperactivity in PTSD patients in relation to matched comparison subjects (left thalamus: p=0.004; right thalamus: p=0.06; data not shown). Thalamic hypoactivity was also more commonly seen in PTSD than either social anxiety disorder or specific phobia
we observed significant negative coactivation between the dorsal or rostral anterior cingulate cortex and the amygdala or insula, indicating that hypoactivation of frontal regions was associated with hyperactivation in limbic and perilimbic regions (see
Hypoactivation of the dorsal amygdala, containing the central nucleus, may be relevant for the autonomic blunting associated with emotional numbing or dissociation in PTSD (74). Hypoactivation of the anterior hippocampus may be important for both declarative memory (75) and regulation of the hypothalamic-pituitary-adrenal axis (76, 77), both of which are perturbed in PTSD (78,79)
amygdala activity strongly habituates to repeated presentation of emotional stimuli (48, 82, 83), which may result in below-baseline levels of activity (48, 82). Although such habituation favors elimination of patient-comparison subject differences rather than hypoactivity (84), it suggests that greater attention should be paid to the time course of amygdala effects
while each of the three disorders involves an excessive fear component, PTSD is a more complex disorder in which the fear part per se is only one element. Thus, PTSD symptoms may be more attributable to dysfunctional emotion regulation systems, whereas the symptoms of social anxiety disorder and specific phobia may be more readily described as intense states of fear
emotion regulation has been investigated using tasks that instruct subjects to deliberately decrease their emotional responses using distraction, reappraisal, suppression, or detachment strategies (95–100). These studies consistently point to involvement of the lateral prefrontal cortex, a locus of executive control for nonemotional stimuli (99, 101). A different picture emerges when one considers “reflexive” forms of emotion regulation, in which the generation and/or modulation of emotion is based on an individual’s expectations about stimuli but without any self-reflective focus on the emotions themselves or the explicit goal of regulating them
Etkin et al. (102) recently found that monitoring of emotional conflict was associated with dorsomedial prefrontal activation, whereas resolution of emotional conflict was associated with rostral anterior cingulate cortex increases and amygdala decreases, consistent with its top-down inhibition. Extinction of conditioned fear also involves increased activity in the rostral anterior cingulate cortex and ventromedial prefrontal cortex and decreased activity in the amygdala (54). Likewise, rostral anterior cingulate cortex activation and amygdala decreases have been observed during placebo anxiolysis (103), and placebo-induced increases in mu-opioid activity have been found in the rostral anterior cingulate cortex, the ventromedial prefrontal cortex, and the amygdala while the subject was experiencing pain (104). Based on these data, we have previously proposed that emotional control processes mediated by the rostral anterior cingulate cortex/ventromedial prefrontal cortex may reflect an individual’s emotional coping or resilience mechanisms (102), which normally function in absence of explicit task instructions to regulate emotion
Our meta-analysis demonstrated robust hypoactivations in PTSD in the rostral anterior cingulate cortex and ventromedial prefrontal cortex but failed to show alterations in the lateral prefrontal cortex. Thus, we propose that hypoactivation of the rostral anterior cingulate cortex and ventromedial prefrontal cortex in patients with PTSD reflects a deficit in reflexive emotion regulation processes occurring in the absence of self-reflection about emotion or deliberate attempts at emotional control and is reflected clinically in emotional dysregulation symptoms and anxiety generalization. A reflexive emotion regulation deficit may thus encompass and extend beyond a fear extinction deficit in PTSD, as has been proposed previously (42,105). The work on the rostral anterior cingulate cortex and the ventromedial prefrontal cortex in humans parallels animal data showing that ventromedial prefrontal cortex lesions in rats impair the ability of these animals to extinguish learned fear (106). Electrical stimulation of this region, which has direct inhibitory projections to the amygdala (107, 108), facilitates fear extinction (109). Although the ventromedial prefrontal cortex in rodents is not an exact homologue of the human rostral anterior cingulate cortex and the ventromedial prefrontal cortex, the shared roles of these areas in fear extinction suggest some degree of analogous function (105) and support a translational approach to anxiety
Unlike the regulatory roles proposed for the rostral anterior cingulate cortex and ventromedial prefrontal cortex, activity in the dorsomedial prefrontal cortex and adjacent dorsal anterior cingulate cortex seems to relate to emotional experience (59) or awareness (110). These regions are activated by emotional conflict (102), track levels of emotional arousal (111), correlate with autonomic activity (112, 113), and respond in anticipation of an aversive event (96), among related functions. Thus, inasmuch as the dorsomedial prefrontal cortex and dorsal anterior cingulate cortex function in the experience of negative emotion, hypoactivation of these regions in PTSD may be related to a decrease in the experience or impact of negative emotion. Because these regions may help recruit rostral anterior cingulate cortex emotion regulation mechanisms (102), dysfunction of the dorsomedial prefrontal cortex and dorsal anterior cingulate cortex may indirectly further contribute to emotional dysregulation in PTSD. Likewise, hypoactivation of the thalamus may relate to decreased processing of sensory information and thereby decreased experience of negative emotion, as suggested previously (40, 72), although the thalamus also plays diverse roles in cortical-cortical interactions
control over the stressor promotes resilience (114–116)
perceived controllability during a trauma is related to the severity of subsequent PTSD symptoms (117)
Eubanks L, Rogers C, Beuscher A, Koob G, Olson A, Dickerson T, Janda K. 2006. “A molecular link between the active component of marijuana and Alzheimer’s disease pathology” Mol Pharm 3(6): 773-777
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2562334/
Δ9-tetrahydrocannabinol (THC), competitively inhibits the enzyme acetylcholinesterase (AChE) as well as prevents AChE-induced amyloid β-peptide (Aβ) aggregation, the key pathological marker of Alzheimer’s disease
Ewijk H, Heslenfeld D, Zwiers M, Buitelaar J, Oosterlaan J. 2012. “Diffusion tensor imaging in attention deficit/hyperactivity disorder: A systematic review and meta-analysis” Neuroscience and Biobehavioural Reviews 36: 1093-1106
http://www.ncbi.nlm.nih.gov/pubmed/22305957 in dropbox
Ewijk H, Heslenfeld D, Zwiers M, Faraone S, Luman M, Hartman C, Hoekstra P, Franke B, Buitelaar J, Oosterlaan J. 2014. “Different Mechanisms of White Matter Abnormalities in Attention-Deficit/Hyperactivity Disorder: A Diffusion Tensor Imaging Study” Journal of the American Academy of Child & Adolescent Psychiatry 53(7): 790-799
http://www.ncbi.nlm.nih.gov/pubmed/24954828 in dropbox
Fachner J. 2003. “Jazz, improvisation and a social pharmacology of music.” Music Therapy Today 4(3)
Fagan S, Campbell V. 2013. “The influence of cannabinoids on generic traits of neurodegeneration.” British Journal of Pharmacology 2014: 171; 1347-1360
http://onlinelibrary.wiley.com/doi/10.1111/bph.12492/full in dropbox
“mechanisms that underlie their neurodegenerative components have been elucidated; namely neuroinflammation, excitotoxicity, mitochondrial dysfunction and reduced trophic support… application of phytocannabinoids and synthetic cannabinoids have been investigated. Signalling from the CB1 and CB2 receptors are known to be involved in the regulation of Ca 2+ homeostasis, mitochondrial function, trophic support and inflammatory status, respectively, while other receptors gated by cannabinoids such as PPAR γ, are gaining interest in their anti-inflammatory properties. Through multiple lines of evidence, this evolutionarily conserved neurosignalling system has shown neuroprotective capabilities and is therefore a potential target for neurodegenerative disorders”
Fanous A, Gardner C, Walsh D, Kendler K. 2001. “Relationship between positive and negative symptoms of schizophrenia and schizotypal symptoms in nonpsychotic relatives” Arch Gen Psychiatry 58(7): 669-673
http://archpsyc.jamanetwork.com/article.aspx?articleid=481805
Positive and negative symptoms in schizophrenia predict corresponding schizotypal symptoms in relatives. This provides evidence that these schizophrenic symptom factors (1) are etiologically distinct from each other and (2) occur on an etiological continuum with their personality-based counterparts.
Positive schizotypy comprises ideas of reference, illusions, and magical thinking, while negative schizotypy includes poor rapport, aloofness, and guardedness.21
Overall, negative symptoms had statistically significant relationships with more schizotypy factors than did positive symptoms. This coheres with the general notion that negative symptoms have greater familial, and possibly genetic, bases than do positive symptoms. Evidence suggesting this is their association with greater family history,37 worse premorbid functioning,38– 40 and greater longitudinal stability.41– 43 Furthermore, the phenomenological resemblance between positive schizotypal symptoms such as magical thinking and illusions on one hand, and positive schizophrenic symptoms on the other, appears to be less than that between negative symptoms of schizotypy21 and schizophrenia.14
The effect sizes (regression slopes) of the analyses did not change substantially when the definition of proband affection was broadened to include nonschizophrenic psychotic disorders, while significance levels increased substantially. This supports the spectrum concept of schizophrenia—that several disorders share with schizophrenia the same underlying liability.47– 50
Fassbender C, Zhang H, Buzy W, Cortes C, Mizuiri D, Beckett L, Schweitzer J. 2009. “A lack of default network suppression is linked to increased distractibility in ADHD” Brain Research 1273: 114-128
http://www.ncbi.nlm.nih.gov/pubmed/19281801 in dropbox
greater variability was associated with a failur e to deactivate vent romedial PFC with increasing task difficulty. T-tes ts on brain activation betwee n participan ts wit h ADHD with low versus high IIV implicated a similar region so that high variability was associated w ith greater acti vity in this region. These data provide support for the theory that incre ased distractibility in at least
some participants with ADHD may be due to an inability to sufficiently suppress activity in the defaul t attention network in respons e to increasing task difficulty
ADHD is a disorder typically character-ized by high degrees of intra-individual variability (IIV) in reaction time (RT) (e.g., Castellanos et al., 2005; Leth-Steensen et al., 2000)
The ADHD subjects evidenced decreases in connectiv-ity between the posterior cingulate/precuneus and task-active regions such as dorsal ACC but also other default network regions such as ventromedial PFC
a reduction in power in very low frequency oscillations electrodes consistent with default network regions was found in young adults with greater numbers of ADHD symptoms, particularly symptoms relating to inattention (Helps et al., 2008). Low frequency oscillations in resting state fMRI data have previously been found to reflect interactions in the default attention network
(De Luca et al., 2006)
The default attention network is a group of regions located along the brain’s medial wall, which are active during non-cognitively demanding paradigms and have been associated with task-irrelevant thought processes, mind wandering, and attention to the outside environment or one’s own mental state (Gusnard et al., 2001b; Shulman et al., 1997)
Inability to sufficiently suppress activity in the default network during a cognitively demanding paradigm may lead to less focus on the task at hand and more vulnerability to distraction from the external environment or one’s own internal non-task-related thoughts ( Weissman et al., 2006)
normally adaptive state of periodically attend-ing to potential novel events in the environment (subserved by the default attention network) can become maladaptive and interfere with ongoing task-related processes… if activation in the default network surpasses a particular threshold, there is a potential for competition with task-specific attention processes, leaving the individual open to attention lapses and decrements in performance. Castellanos et al. (Sonuga-Barke and Castellanos, 2007 ; Castellanos et al., 2008) purport this as a m odel for attention deficits in ADHD with inattention stemmin g f rom a n i mbala nce between suppressing the default network and activating task-appro-priate regions
However, contrary to Castellanos et al. (Castellanos et al., 2008; Uddin et al., 2008 ) finding of dysfunction in the posterior cingulate in ADHD, we found no differences in patterns of deactivation in this region between ADHD and HC children in either the simpler AT task or the more cognitively dema nding VS AT. This held tr ue fo r b ot h t he post erior cingulate region defined by the HC group and that defined by the ADHD group. Group differences were only found in medial PFC. As can be seen in Fig. 2, although both groups tended to increase deactivation from AT to VSAT in three regions in medial PFC/ACC, the ADHD group failed to suppress activity in these regions to the same degree as their HC peers
those with greater RT variability d uring the VSAT also tend ed to di splay l ess deactivation in the ventromedial PFC during the working memory task
study examining default network activity revealed attenuated levels of activation in medial PFC, ACC and poster-ior cingulate in children compared to adults (Marsh et al., 2006). This finding was interpreted as being due to greater auto-maticity of performance on the baseline task in adults leading to an incr ease in task-irr elevan t t hou ghts co mpar ed t o children
Although Castellanos et al. (2008)suggest that the region of the default network incorporating posterior cingulate extend-ing into precuneus may be a core of dysfunction in ADHD participants responsible for increased variability, we found no suggestion of any difference between ADHD and HC partici-pants in this region
In children, proximal regions tend to be interconnected more than in adults, whereas distal regions, such as medial PFC/rostral ACC and posterior cingulate, tend to be less interconnected (Fair et al., 2007 ). Default network structure in adolescents is thought to lie somewhere between that of children and adults, with shorter range connections being progressively broken and longer range connections being formed (Fair et al., 2007)
it could be argued that only five of the children with ADHD are driving our results… It may be that the default network hypothesis is applicable to at least a large subgroup of individuals with ADHD
Felitti V, Anda R, Nordenberg D, Williamson D, Spitz A, Edwards V, Koss M, Marks J. 1998. “Relationship of Childhood Abuse and Household Dysfunction to Many of the Leading Causes of Death in Adults: The Adverse Childhood Experiences (ACE) Study” American Journal of Preventative Medicine 14:245-258
http://www.ncbi.nlm.nih.gov/pubmed/9635069 in dropbox
“smoking, which is medically and socially viewed as a ‘problem’ may, from the perspective of the user, represent an effective immediate solution that leads to chronic use. … incomplete understanding of the possible benefits of health risk behaviors leads them to be viewed as irrational and having solely negative consequences”
Fattore L, Melis M, Fadda P, Pistis M, Fratta W. 2010. “The endocannabinoid system and nondrug rewarding behaviours” Experimental Neurology 224: 23-36
http://www.ncbi.nlm.nih.gov/pubmed/20353776 in dropbox
rewarding behaviours activate the same brain circuits that mediate the postiive reinforcing effects of drugs
we focus first on the role of the endocannabinoid system in the modulation of neural activity and synaptic functions in brain regions that are involved in natural and nonnatural rewards (namely, the ventral tegmental area, striatum, amygdala and prefrontal cortex)
marijuana and cannabinoids, in general, are potent modulators of the central mechanisms that underlie not only drug (Fattore et al, 2007a, 2008) but also nondrug rewarding behaviours
Felitti V, Anda R. 2009. “The relationship of adverse childhood experiences to adult medical disease, psychiatric disorders and sexual behaviour: implications for healthcare” chapter in The Hidden Epidemic: The Impact of Early Life Trauma on Health and Disease Cambridge University Press. ISBN-13: 978-0521880268
http://www.acestudy.org/yahoo_site_admin/assets/docs/LaniusVermetten_FINAL_8-26-09.12892303.pdf in dropbox
1/3 had 0 ACEs, 1/6 had 4 ACEs, 1/9 had 5 ACEs 54% of current depression can be attribute to ACEs; males with an 6 ACEs 4,600% more likely to inject drugs than 0 ACE male
“It’s hard to get enough of something that almost works” “the attempted solution rather than the core problem is being addressed”
strong relationship of ACE Score to coronary disease, after correcting for all the conventional risk factors like smoking, cholesterol
Fellner J, Walsh S, Roth K, Smart, M. 2000. “Punishment and Prejudice: Racial Disparities in the War on Drugs” Human Rights Watch 12(2)
Fennig S, Horesh N, Aloni D, Apter A, Weizman A. 2005. “Life events and suicidality in adolescents with schizophrenia” Eur Child Adolesc Psychiatry 14(8): 454-60
http://www.ncbi.nlm.nih.gov/pubmed/16341502
number of negative events differentiates from non-schizophrenics, but not suicidal from non-suicidal. Ratio negative to other life events does: suicidal 74%, nonsuicidal 57%
Fergusson D, Horwood L. 1997. “Early onset cannabis use and psychosocial adjustment in young adults.” 92(3): 279-96
http://www.ncbi.nlm.nih.gov/pubmed/9219390/ in dropbox
those who used by age 16, esp frequent users more likely to come from socially disadvantaged backgrounds, experienced family adversity as children, formed affiliations with substance using or delinquent peers in adolescence, reported poorer attachments with parents, had more adjustment problems by the age of 16. Early onset users had significantly higher rates of later substance use, juvenile offending, mental health problems, unemployment and school dropout
Fergusson D, Swain-Campbell N, Horwood L. 2003. “Arrests and convictions for cannabis related offences in a New Zealand birth cohort” Drug Alcohol Depend 70(1):53-63
http://www.ncbi.nlm.nih.gov/pubmed/12681525
Over a quarter of those using cannabis more than 400 times were arrested or convicted (5.1% of total users, who are more than two thirds of the cohort). Males and Maori overrepresented. Of those arrested/convicted, 95% either increased use or continued with same level of use subsequent to arrest.
Fergusson D, Horwood L, Ridder E. 2005. “Tests of causal linkages between cannabis use and psychotic symptoms” Addiction 100: 354-366
http://www.ncbi.nlm.nih.gov/pubmed/15733249 in dropbox
Fernández-López D, Martinez-Orgado J, Nunez E, Romero J, Lorenzo P, Moro M, Lizasoain I. 2006. “Characterization of the neuroprotective effect of the cannabinoid agonist WIN-55212 in an in vitro model of hypoxic-ischemic brain damage in newborn rats” Pediatr Res 60(2): 169-73
http://www.ncbi.nlm.nih.gov/pubmed/16864698/
WIN55212 afforded robust neuroprotection in the forebrain slices exposed to OGD, by acting on glutamatergic excitotoxicity, TNF-alpha release, and iNOS expression; this neuroprotective effect seemed to be mediated by CB1 and CB2 receptors
Ferrer B, Asbrock N, Kathuria S, Piomelli D, Giuffrida A. 2003. “Effects of levodopa on endocannabinoid levels in rat basal ganglia: implications for the treatment of levodopa-induced dyskinesias” Eur J Neurosci 18(6): 1607-14
http://www.ncbi.nlm.nih.gov/pubmed/14511339
In intact animals, systemic administration of levodopa increased anandamide concentrations throughout the basal ganglia via activation of dopamine D1/D2 receptors… a deficiency in endocannabinoid transmission may contribute to levodopa-induced dyskinesias and that these complications may be alleviated by activation of CB1 receptors
Filbey F, Aslan S, Calhoun V, Spence J, Damaraju E, Caprihan A, Segall J. 2014. “Long-term effects of marijuana use on the brain” PNAS 111(47): 16913-16918
http://www.pnas.org/content/111/47/16913.full
findings remain inconclusive
[MV: the finding of decreased FA from chronic use was mainly driven by the two longest using subjects]
The results showed that compared with controls, marijuana users had significantly less bilateral orbitofrontal gyri volume, higher functional connectivity in the orbitofrontal cortex (OFC) network, and higher structural connectivity in tracts that innervate the OFC (forceps minor) as measured by fractional anisotropy (FA). Increased OFC functional connectivity in marijuana users was associated with earlier age of onset
Although some have reported decreases in regional brain volume such as in the hippocampus, orbitofrontal cortex, amygdala, and striatum (7⇓⇓⇓⇓–12), others have reported increases in amygdala, nucleus accumbens, and cerebellar volumes in chronic marijuana users (13⇓–15). However, others have reported no observable difference in global or regional gray or white matter volumes in chronic marijuana users (16, 17)
110 participants consisting of 62 nonusing controls and 48 marijuana users… included if they currently use marijuana regularly (at least four times per week) over the last 6 mo (confirmed via positive THC-COOH urinalysis) [from table 1, half of users were considered dependent]
the IQ of the marijuana users was significantly lower than the control group (P < 0.05)
quantitatively, marijuana users had significantly higher connectivity in all four nodes (i.e., bilateral OFC and bilateral temporal lobe) compared to the control group
earlier age of onset leads to higher functional connectivity of the bilateral OFC. Finally, there was an inverse correlation between the left temporal lobe functional connectivity and problems related with marijuana use, such that the greater the functional connectivity of the left temporal cortex to bilateral OFC, the lower the total score on MPS
inverse relationship between bilateral OFC gray matter volume and problems related to marijuana use. That is, the lower the OFC gray matter volume in these participants, the higher their MPS total scores
We did not find that the causal variable (i.e., marijuana use) was significantly correlated with the mediator variable (i.e., OFC gray matter volume, OFC/temporal functional connectivity, and FA/RD of forceps minor) and outcome variable (i.e., IQ). We, therefore, suggest that the path from marijuana use to neural abnormalities to decreases in IQ is more complex and, perhaps, include other mediators such as environmental (i.e., age of onset) and/or genetic factors.
a prospective study also found that smaller OFC volumes at 12 y of age predicted initiation of marijuana use at 16 y of age (30)
these OFC abnormalities may reflect preexisting pathophysiology related to vulnerability to marijuana abuse and dependence
Greater FA has been suggested to reflect better myelination and/or intact axons (42).
FA difference between the groups in the forceps minor was driven by lower RD, suggesting greater myelination in the marijuana users
Greater FA has also been reported in alcohol users (44, 45), which was posited to reflect a premorbid vulnerability for accelerated PFC myelin maturation in those at risk for alcohol use disorders
DTI is sensitive to increased tissue water resulting in decreased FA as a result of inflammation; therefore, it is possible that any antiinflammatory effects of cannabis would lead to greater FA. Lastly, it is also possible that the effects of cannabis (i.e., CBD) may be beneficial to white matter in terms of regulation of mitochondrial activity, antioxidant processes, and modulation of clearance processes that protect neurons on the molecular level (46)
greater functional connectivity was associated with earlier onset of regular use, whereas chronic marijuana use showed lower structural connectivity (i.e., FA)
Fink M. 1976. “Effects of acute and chronic inhalation of hashish, marijuana, and delta 9-tetrahydrocannabinol on brain electrical activity in man: evidence for tissue tolerance” Ann N Y Acad Sci 282: 387:98
http://www.ncbi.nlm.nih.gov/pubmed/1071390/
The EEG effects of enhanced alpha activity, decreased beta activity, and decreased mean frequency are dose dependent, both in intensity and in duration. The behavioral measures, particularly self-ratings of euphoria (“high” or “mastura”) and heart rate, are also dose dependent and interrelated with the EEG measures. Tolerance to the cerebral, heart rate, and behavioral effects of repeated inhalations of delta 9-THC are demonstrable in chronic users and also in short-term (21-day) experimental administration studies… No differences in incidence of abnormal EEG records were found between long-term heavy hashish users and controls. Taken in conjunction with the neurologic, echo-EEG, and psychopathologic data in these men, we failed to define a syndrome of persistent brain damage as a result of hashish use, although persistent adaptational changes are noted in the development of tissue tolerance.
Finkelhor D, Hotaling G, Lewis I, Smith C. 1990. “Sexual abuse in a national survey of adult men and women: prevalence, characteristics and risk factors.” Child Abuse Negl 14:19-28
http://www.ncbi.nlm.nih.gov/pubmed/2310970
27% of women and 16% of men had been sexually abused
Finkelhor D, Ormrod R, Turner H. 2009. “Lifetime assessment of poly-victimization in a national sample of children and youth” Child Abuse & Neglect 33 403-411
http://unh.edu/ccrc/pdf/CV176.pdf
Nearly 80% of the children and youth victimized; mean lifetime victimizations 3.7, median 2.6. Number highly predictive of current distress symptoms. Best linear prediction weights: child maltreatment 4, sexual assault 3, other victimizations 1. propose classifying poly-victims as those 10% with the highest scores for each age threshold
Fishbein M, Gov S, Assaf F, Gafni M, Keren O, Sarne Y. 2012. “Long-term behavioral and biochemical effects of an ultra-low dose of Δ9-tetrahydrocannabinol (THC): neuroprotection and ERK signaling” Exp Brain Res 221(4): 437-48
http://www.ncbi.nlm.nih.gov/pubmed/22821081
We have previously reported that a single injection of an ultra-low dose of delta-9-tetrahydrocannabinol (THC; the psychoactive ingredient of marijuana) protected the brain from pentylenentetrazole (PTZ)-induced cognitive deficits when applied 1-7 days before or 1-3 days after the insult… we expanded the protective profile of THC by showing that it protected mice from cognitive deficits that were induced by a variety of other neuronal insults, including pentobarbital-induced deep anesthesia, repeated treatment with 3,4 methylenedioxymethamphetamine (MDMA; “ecstasy”) and exposure to carbon monoxide. The protective effect of THC lasted for at least 7 week… long-lasting (7 weeks) modifications of extracellular signal-regulated kinase (ERK) activity in the hippocampus, frontal cortex and cerebellum of the mice. The alterations in ERK activity paralleled changes in its activating enzyme MEK and its inactivating enzyme MKP-1. Furthermore, a single treatment with the low dose of THC elevated the level of pCREB (phosphorylated cAMP response element-binding protein) in the hippocampus and the level of BDNF (brain-derived neurotrophic factor) in the frontal cortex. These long-lasting effects indicate that a single treatment with an ultra-low dose of THC can modify brain plasticity and induce long-term behavioral and developmental effects in the brain
Fisher H, Jones P, Fearon P, Craig T, Dazzan P, Morgan K, Hutchinson G, Doody G, McGuffin P, Leff J, Murray R, Morgan C. 2010. “The varying impact of type, timing and frequency of exposure to childhood adversity on its association with adult psychotic disorder” Psychological Medicine 40: 1967-1978
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3272393/ in dropbox
Psychosis cases were three times more likely to report severe physical abuse from mother that commenced prior to 12 years of age, even after adjustment for other significant forms of adversity and demographic confounders
Associations with maternal neglect and childhood sexual abuse disappeared after adjusting for maternal physical abuse and antipathy. Paternal maltreatment and other forms of adversity were not associated with psychosis nor was there evidence of a dose-response effect
Given the hypothesised key role of mother-child attachment bonds in psychological wellbeing (Bowlby, 1951, 1977), evidence from cohort studies of mothers’ perceptions and care of the child being related to adult schizophrenia (Jones et al., 1994; McNeil et al., 2009; Myhrman et al., 1996), and the greater absence of fathers in the childhoods of individuals with psychosis (Agid et al., 1999; Morgan et al., 2007) it seems plausible to hypothesise that abuse perpetrated by mothers will also have a greater association with psychotic disorders
the vast majority of psychosis cases (86%) and controls (93%) in this study did not report maternal physical or emotional abuse
If comparisons are restricted to the largest previous study of first-episode psychosis patients (Neria et al., 2002), then the rates reported in the current study are slightly lower for physical abuse (21.6% vs. 14.0-15.2%) but fairly similar for sexual abuse (15.0% vs. 18.2%)
the prevalence of physical and sexual abuse reported by healthy controls in this study (5.4-12.3% and 13.1%, respectively) is similar to that found in a recent UK general population survey (May-Chahal & Cawson, 2005: 7-14% physical, 11% sexual)
it is possible that previous associations between sexual abuse and psychosis may have been inflated by overlap with physical abuse
Higher rates of psychotic symptoms have also been found in those exposed to emotional forms of abuse (Colins et al., 2009; Sommer et al., 2008)
the importance of the mother in the psychological development of the child has long been advocated as she is usually the main attachment figure in infancy (Bowlby, 1951, 1977)
it is also plausible that stronger effects in childhood are just a proxy for more prolonged abuse or higher risk of exposure to other adverse experiences
other potentially predictive forms of childhood adversity were not measured, such as bullying, psychological abuse, domestic violence and family discord (see Bebbington et al., 2004; Campbell & Morrison, 2007; Kelleher et al., 2008; Lataster et al., 2006; Rosenberg et al., 2007)
Fisher H, Craig T, Fearon P, Morgan K, Dazzan P, Lappin J, Hutchinson G, Doody G, Jones P, McGuffin P, Murray R, Leff J, Morgan C. 2011. “Reliability and comparability of psychosis patients’ retrospective reports of childhood abuse” Schizophrenia Bulletin 37(3): 546-553
http://schizophreniabulletin.oxfordjournals.org/content/37/3/546.short in dropbox
high levels of concurrent validity were demonstrated with the Parental Bonding Instrument were demonstrated with the Parental Bonding Instrument (antipathy:rs = 0.350–0.737, P < .001; neglect:rs = 0.688– 0.715, P < .001), and good convergent validity was shown with clinical case notes (sexual abuse: k 5 0.526, P < .001; physical abuse: k = 0.394, P < .001). Psychosis patients’ reports were also reasonably stable over a 7-year period (sexual abuse: k = 0.590, P < .01; physical abuse: k = 0.634, P < .001; antipathy: k = 0.492, P < .01; neglect: k = 0.432, P < .05). Additionally, their reports of childhood abuse were not associated with current severity of psychotic symptoms (sexual abuse: U = 1768.5, P = .998; physical abuse: U = 2167.5, P = .815; antipathy: U = 2216.5, P = .988; neglect: U = 1906.0, P 5 .835) or depressed mood (sexual abuse: x2 = 0.634, P 5 .277; physical abuse: x2 = 0.159, P = .419; antipathy: x2 = 0.868, P = .229; neglect: x2 = 0.639, P 5 .274). These findings provide justification for the use in future studies of retrospective reports of childhood abuse obtained from individuals with psychotic disorders
Flinn M, Nepomnaschy P, Muehlenbein M, Ponzi D. 2011. “Evolutionary fuctions of early social modulation of hypothalamic-pituitary-adrenal axis development in humans.” Neurosci Biobehav Rev 35(7): 1611-29
http://www.ncbi.nlm.nih.gov/pubmed/21251923
difficult family environments and traumatic social events are associated with temporal elevations of cortisol, suppressed reproductive functioning and elevated morbidity. The long-term effects of traumatic early experiences on cortisol profiles are complex and indicate domain-specific effects, with normal recovery from physical stressors, but some heightened response to negative-affect social challenge
Folsom D, Hawthorne W, Lindamer L, Gilmer T, Bailey A, Golshan S, Garcia P, Unützer J, Hough R, Jeste D. 2005. “Prevalence and risk factors for homelessness and utilization of mental health services among 10,340 patients with serious mental illness in a large public mental health system” Am J Psychiatry 162(2): 370-6
http://www.ncbi.nlm.nih.gov/pubmed/15677603
The prevalence of homelessness was 15%. Homelessness was associated with male gender, African American ethnicity, presence of a substance use disorder, lack of Medicaid, a diagnosis of schizophrenia or bipolar disorder, and poorer functioning. Latinos and Asian Americans were less likely to be homeless. Homeless patients used more inpatient and emergency-type services and fewer outpatient-type services
Ford J, Hawke J, Alessi S, Ledgerwood D, Petry N. 2007 “Psychological trauma and PTSD symptoms as predictors of substance dependence treatment outcomes” Behav Res Ther 45(10): 2417-31
http://www.ncbi.nlm.nih.gov/pubmed/17531193
affect dysregulation, dissociation, and disturbed interpersonal functioning; C-PTSD predicts poor outcome, PTSD predicts improved outcome
Ford J, Elhai J, Connor D, Frueh B. 2010. “Poly-victimization and risk of posttraumatic, depressive, and substance use disorders and involvement in delinquency in a national sample of adolescents” J Adolesc Health 46(6): 545-52
http://www.ncbi.nlm.nih.gov/pubmed/20472211
poly-victimized youths’ risk of delinquency cannot be fully accounted for by posttraumatic stress disorder, depression, or substance use problems
Di Forti M, Marconi A, Carra E, Fraietta S, Trotta A, Bonomo M, Bianconi F, Gardner-Sood P, O’Conner J, Russo M, Stilo S, Marques T, Mondelli V, Dazzan P, Pariante C, David A, Gaughran F, Atakan Z, Iyegbe C, Powell J, Morgan C, Lynskey M, Murray R. 2015. “Proportion of patients in south London with first-episode psychosis attributable to use of high potency cannabis: a case control study”
http://www.thelancet.com/pb/assets/raw/Lancet/pdfs/14TLP0454_Di%20Forti.pdf in dropbox
Study uses loaded, inaccurate and stigmatising terminology such as “skunk-like” to refer to good quality bud (skunk is one breed among thousands, but British media has seized on it because it sounds uglier than, say, “girl scout cookies” or “tickle kush”) and ignores the real reasons why schizophrenics use marijuana, rejecting self-medication on the grounds that it doesn’t reduce positive symptoms, when it’s in fact used to reduce negative symptoms. A bad-faith propaganda piece from King’s College London, which nevertheless contains some interesting data
Patients with first-episode psychosis were no more likely than were controls to report a lifetime history of ever having used cannabis, but were more likely to use cannabis every day and to mostly use high-potency (skunk-like) cannabis
among cannabis users, the mean duration of use did not differ between patients with first-episode psychosis and controls (table 2). On average, both groups started using cannabis in their mid-teens
individuals who had ever used cannabis were not at increased risk of psychotic disorder compared with those who had never used cannabis (n=775) Individuals who started using cannabis at ages younger than 15 years had modestly, but significantly, increased risk of psychotic disorders compared with those who never used cannabis (table 3). People who used cannabis or skunk every day were both roughly three times more likely to have a diagnosis of a psychotic disorder than were those who never used cannabis (table 3)
Individuals who mostly used low-potency (hash-like) cannabis occasionally (p=0·493), at weekends (p=0·102), or daily (p=0·626) had no increased likelihood of psychotic disorders compared with those who never used cannabis (figure 2)
If causality is assumed, this finding suggests that skunk alone was responsible for the largest proportion of new cases (24%) of psychotic disorder in the study population, an effect driven by its high prevalence among patients with first-episode psychosis who used cannabis (218 [53%] of 410 patients)
The presence of cannabidiol might explain our results, which showed that hash users do not have any increase in risk of psychotic disorders compared with non-users, irrespective of their frequency of use
South London has one of the highest recorded incidence rates of psychosis in the UK. Boydell and colleagues showed that the incidence of schizophrenia had doubled since 1965, and that one possible contribution to this was the increase in cannabis use among individuals who developed schizophrenia. In the present study, we identified an increased estimate for the PAF accounted for by cannabis (24%) compared with previous studies, which reported PAFs of 6·2% in Germany, 8% in New Zealand, and 13·3% in Holland. This finding could be caused by, not only the greater use of cannabis, but also the greater use of high-potency (skunk-like) cannabis in south London than in these other countries in earlier periods [so the zero increase from any cannabis use, which can be made to look like 24% increase from high-THC cannabis use because schizophrenics prefer high THC cannabis because it more effectively treats negative symptoms, is the causal factor explaining why the incidence DOUBLED? Psychiatrists at King’s College London evidently are not strong on arithmetic]
That people who already have prodromal symptoms would choose a type of cannabis that is high in THC and has little cannabidiol (such as skunk), which might exacerbate their symptoms, rather than a cannabidiol-containing type (such as hash), would seem counterintuitive [if you ignore why they use it]
urgent need to develop public education to inform young people about the risks of high-potency cannabis
Foti D, Kotov R, Guey L, Bromet E. 2010. “Cannabis use and the course of schizophrenia: 10-year follow-up after first hospitalization” Psychiatryonline 167(8): 987-993
http://psychiatryonline.org/doi/full/10.1176/appi.ajp.2010.09020189
authors assessed 229 patients with a schizophrenia spectrum disorder five times: during the first admission and 6 months, 2 years, 4 years, and 10 years later
lifetime rate of cannabis use was 66.2%, and survival analysis revealed that lifetime use was associated with an earlier onset of psychosis. The rates of current use ranged from 10% to 18% across assessments
Mixed-effects logistic regression revealed that changes in cannabis use were associated with changes in psychotic symptoms over time even after gender, age, socioeconomic status, other drug use, antipsychotic medication use, and other symptoms were controlled for. Structural equation modeling indicated that the association with psychotic symptoms was bidirectional
A recent review of 53 treatment studies found that the average 12-month prevalence of cannabis use was 29.2% among patients with psychosis (1), compared with 4.0% in the general U.S. population (2)
Some individuals with schizophrenia report using cannabis to relieve symptoms or medication side effects (11 – Dixon). Also, some cross-sectional studies have reported that cannabis use is associated with less severe negative and disorganized symptoms (12 – Bersani)
On the other hand, other cross-sectional data indicate that cannabis users have more severe psychotic symptoms than nonusers (13, 14)
lower severity of psychotic symptoms predicted cessation of cannabis use, whereas higher severity was associated with an increased likelihood of use at the next assessment. Likewise, cannabis use predicted an increase in severity of psychotic symptoms
we cannot rule out the possibility that some individuals used cannabis to obtain relief from disorganized symptoms (11, 12), but in this sample a more robust association was found with psychotic symptoms. This suggests that individuals with schizophrenia who use cannabis are not more severely ill overall but suffer specifically from more severe psychotic symptoms.
Frank M, Baratta M, Sprunger D, Watkins L, Maier S. 2007. “Microglia serve as a neuroimmune substrate for stress-induced potentiation of CNS pro-inflammatory cytokine responses” Brain Behav Immun 21(1): 47-59
http://www.ncbi.nlm.nih.gov/pubmed/16647243/
microglia are the primary immune effector cell in the CNS… The present results suggest that stress can activate microglia, thereby sensitizing the pro-inflammatory reactivity of microglia to immunogenic stimuli
Franklin A, Parmentier-Batteur S, Walter L, Greenberg D, Stella N. 2003. “Palmitoylethanolamide increases after focal cerebral ischemia and potentiates microglial cell motility” J Neurosci 23(21): 7767-75
http://www.ncbi.nlm.nih.gov/pubmed/12944505
FCI greatly increases palmitoylethanolamide (PEA), only moderately increases anandamide [arachidonylethanolamide (AEA)], and does not affect 2-arachidonoylglycerol levels. We also found that PEA potentiates AEA-induced microglial cell migration, without affecting other steps of microglial activation, such as proliferation, particle engulfment, and nitric oxide production
French L, Gray C, Leonard G, Perron M, Pike B, Richer L, Seguin J, Veillette S, Evans J, Artiges E, Banaschewski T, Bokde A, Bromberg U, Bruehl R, Buchel C, Cattrell A, Conrod P, Flor H, Frouin V, Galliant J, Garavan H, Gowland P, Heinz A, Lemaitre H, Martinot J-L, Nees F, Orfanos D, Pangelinan M, Poustka L, Rietschel M, Smolka M, Walter H, Whelan R, Timpson N, Schumann G, Smith G, Pausova Z, Paus T. 2015. “Early cannabis use, polygenic risk score for schizophrenia, and brain maturation in adolescence” JAMA Psychiatry 72(10): 1002-1011
http://archpsyc.jamanetwork.com/article.aspx?articleid=2429551 add to dropbox
initial analysis in 1024 adolescents of both sexes from the Canadian Saguenay Youth Study (SYS) and follow-up in 426 adolescents of both sexes from the IMAGEN Study from 8 European cities and 504 male youth from the Avon Longitudinal Study of Parents and Children (ALSPAC) based in England. A total of 1577 participants (aged 12-21 years; 899 [57.0%] male) had (1) information about cannabis use; (2) imaging studies of the brain; and (3) a polygenic risk score for schizophrenia across 108 genetic loci identified by the Psychiatric Genomics Consortium
Across the 3 samples of 1574 participants, a negative association was observed between cannabis use in early adolescence and cortical thickness in male participants with a high polygenic risk score. This observation was not the case for low-risk male participants or for the low- or high-risk female participants
2011 European School Survey Project on Alco-hol and Other Drugs,3 a mean lifetime prevalence of canna-bis use among high school students aged 15 to 16 years was 17%, with large variations across the 36 participating coun-tries (eg, 19% in Germany, 25% in the United Kingdom, and 39% in France). The 2014 Monitoring the Future survey4 has reported a lifetime prevalence of cannabis use of 35.8% among youth aged 15 to 16 years living in the United States in 2013
steeper slopes of age-related increases in white matter and decreases in (cortical) gray matter in male compared with female adolescents.13,14 These sex differences in the dynamics of brain maturation during adolescence may be of particular importance with regard to vulnerability of the male brain to external factors, such as cannabis exposure, during this period of development
SYS sample 1024 adolescents comes from the Saguenay Lac-Saint-Jean region of Quebec, Canada… first replication sample consisted of 504 male youth recruited from the Avon Longitudinal Study of Parents and Children (ALSPAC)20 based in England… second replication sample consisted of 426 adolescents recruited in 8 European cities in the context of the IMAGEN Study
SYS sample classified adolescents as ever or never used cannabis. In ALSPAC and IMAGEN address the latter question using original data on the number of occasions of cannabis use by 16 years of age
cortical thickness is a useful metric for capturing the cumu-lative effects of various experiential factors on cortical neu-robiological features, especially neuropil (ie, dendrites, glial cells) and capillary densities
we have related regional variations in the group differ-ences (users vs nonusers) in thickness across 34 cortical regions to those in the expression of the cannabinoid receptor 1 gene ( CNR1 [NCBI Entrez Gene 1268]) derived from the Allen Brain Atlas in the same regions.26 This atlas provides postmortem measurements of gene expression obtained in 6 adult brains (1269 cortical samples were used to calculate an aver-age for each of the 34 regions). We used CNR1 expression as a proxy of the cannabinoid type 1 receptor density to evaluate whether the extent of the relationship between cannabis use and cortical thickness varies as a function of this receptor’s density in the cerebral cortex, thus testing for the level of specificity in this relationship
The Schizophrenia Working Group of the Psychiatric Genomics Consortium found that the top decile (based on the top 108 loci) contained about 3 times more cases of schizophrenia than the bottom decile (mean odds ratio across 39 samples, 3.21)
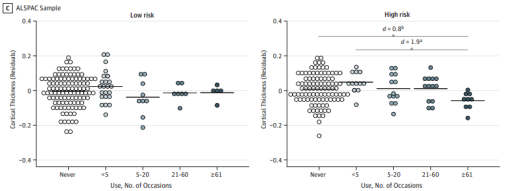
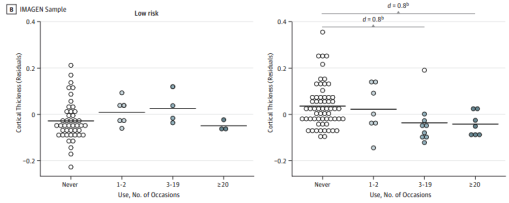
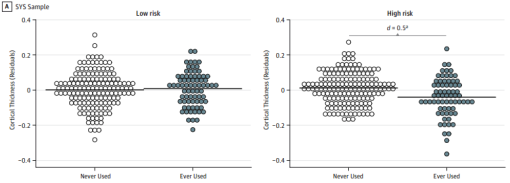
the largest group differences between those who never and ever used cannabis were found in regions that showed high CNR1 expression(eg,entorhinal and anterior cingulate cortex)
Observational studies such as ours cannot attribute causality to the observed relationships. Even the longitudinal de-sign does not rule out the possibility that individuals with a particular developmental trajectory may be more likely to experiment with cannabis rather than the cannabis exposure affecting the trajectory
Unlike the SYS and IMAGEN samples, the high-risk male participants in the ALSPAC sample do not show a difference in cortical thickness between those who never and ever used cannabis; only the high-frequency users do. We can only speculate that, with a given sample size, the association between less frequent cannabis use and cortical thickness is less robust and, therefore, sensitive to other (confounding) effects that may accumulate with age
The overall volume of cortical gray matter and cortical thickness decrease with age in typi-cally developing male adolescents. 13,14 Our longitudinal findngs suggest that cannabis exposure might accelerate such processes, including cortical thinning, in male adolescents with a high polygenic risk score. A profound thinning of cortical gray matter was observed during adolescence in patients with childhood-onset schizophrenia (onset of symptoms by 12 years of age)31,32 and, to a much lesser extent, in their nonpsychotic siblings.33
Rising levels of testosterone during male adolescence and the associated high dynamics in the neurobiological features underlying cortical maturation may represent a risk factor with regard to other external (eg, cannabis) and/or internal eg, genetic risk) perturbations. Furthermore, limited evidence supports the possible effects of testosterone on potentiating the action of cannabinoid type 1 receptor agonists on presynaptic inhibition of excitatory inputs in vitro57 and on transcriptional upregulation of the CNR1 gene.58,59
In this report, the polygenic risk score for schizophrenia calculated with the genome-wide significant SNPs ( P <5×10−8) showed an association with cortical thickness. This association was not in evidence when we calculated the score with the 24 727 nominally significant SNPs ( P < .05)(eTable4 in the Supplement ). Nevertheless, the latter score is superior to the former in predicting liability to schizophrenia.17
we speculate that the moderating in-fluence of cannabis use on the association between the ge-netic risk for schizophrenia and cortical thickness may repre-sent a combination of reduced experience-related brain plasticity taking place on the background of testosterone-associated decreases in cortical gray matter
Fried P, Watkinson B, James D, Gray R. 2002. “Current and former marijuana use: preliminary findings of a longitudinal study of effects on IQ in young adults.” CMAJ 166(7):887-91
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC100921/ in dropbox
IQ difference scores showed decrease of 4.1 points in current heavy users (more than 5 joints/week) ( p < 0.05); gains in IQ for light current users (5.8), former users (3.5) and non-users (2.6)
potentially confounding variables: socioeconomic status (incl family income and parental education); the subject’s education level (years of education); age and sex of the subject; mother’s age at the time of birth; maternal use of cigarettes, marijuana and alcohol during pregnancy; use of tobacco and alcohol and exposure to secondhand marijuana smoke.
no predictive relation with the IQ difference score was found for the self-reported period of marijuana use or the estimated total number of joints smoked
lack of a negative impact among the former heavy users; they had smoked avg 5793 joints over 3.2 years (mean of 37 joints per week); the current heavy users had smoked, on average, an estimated 2386 joints over 3.1 years (mean of 14 joints per week)
Fried P, Watkinson B, Gray R. 2005. “Neurocognitive consequences of marihuana–a comparison with pre-drug performance.” Neurotoxicol Teratol 27(2):231-9
http://www.ncbi.nlm.nih.gov/pubmed/15734274
current regular heavy users did significantly worse than non-users in overall IQ, processing speed, immediate, and delayed memory; former marihuana smokers did not show any cognitive impairments
there is considerable consensus that gross cognitive impairment is not evident in chronic users compared to irregular or non-users following lengthy abstinence periods [13,29,3 6,40,42]
Ottawa Prenatal Prospective Study (OPPS) was initiated in 1978 with the primary objective of examining the effects on offspring of soft drugs used during pregnancy. In this predominantly middle class, low risk sample, approximately 160 children have been administered neuropsychological tests yearly to age 7 and once during each of the 9-12, 13-16, and 17-21 year intervals… in the present study, data from the 9-12 year testing (pre-teen) were used as pre-drug performance measures for the 17-21 year testing (young adult)… of the 152 subjects who have been tested between 17 and 21, 121 were available who had also been tested at 9-12
Additional exclusions were one subject on ritalin, one subject because of amphetamine use, an LSD user, an uncooperative subject, three cases with a positive urinalysis for cannabinoid metabolites coupled with a negative self-report, and one previous regular user who had quit for several months but began regular use a few weeks before testing. The final sample comprised 113 subjects, 50 females and 63 males. None smoking marihuana regularly at the 9-12 year testing. Of the 113 subjects, 10 had tried between 9 and 12: 1 from the control group and 2, 6, and 1 from the current light, current heavy, and former user group, respectively
Potential confounds considered were SES variables (family income, parental education), maternal use of alcohol, cigarettes, and marihuana during pregnancy, age and sex of subject, young adult’s cigarette and alcohol use, and DSM positive criteria for any of the following DSM-IV Axis I disorders: generalized anxiety, major depression, dysthymic disorder, attention deficit/hyperactivity disorder, conduct disorder, oppositional defiant disorder, alcohol dependence and abuse.
Although on average, current and former marihuana useing groups showed more positive DSM criteria than the comparison group, the frequency of positive diagnoses for each of the disorders considered was low
After controlling for pre-use performance, the current heavy use group had lower mean scores in Immediate Memory and General (delayed) Memory as well as the Processing Speed Index, a composite score contributing to overall IQ
former users smoked 2203 joints on average, compared to 1884 in current heavy users and 122 in current light. 9.6 cigarettes per day in heavy current group, and 8.8 maternal joints per week (1.5-2.1 in other groups)
After controlling for pre-drug performance , no relation-ships were found between current marihuana use and working memory [48] , Verbal IQ [47] , the Peabody Picture Vocabulary Test (PPVT) [44] , the Category Test [44] , or the TOVA attention tasks [26]
Processing Speed Index was associated with current heavy marihuana use after, but not before, pre-drug performance control. Conversely, both the PPVT and Category Test were associated with current heavy marihuana use before, but not after, pre-drug performance control
Self-reports of marihuana usage in each of the 7 days previous to testing, when individually correlated with relevant outcomes, did not reveal stroner negative associations with more recent exposure… no significant differences between former heavy and comparison group on any of the cognitive outcomes were noted
former user group started regular use slightly younger (14.3 years) than the current heavy users (15 years), and consumed a total of approximately 5000 joints compared with 2000 for the current users
deficits in visual information proces sing are largely independent of competency in visual attention as demonstrated both by the low correlations between the visual attention task and the Processing Speed Index in the present work as well as with Wechsler ’s report of only low to moderate correlations between several measures of attention and the Visual Processing composite score [49] .
The absence of a negative impact among the former heavy users in the present report is all the more striking as they initiated their regular habits at a younger age than the curren users (14 vs 15 years)… the cognitive impairments among the current heavy users and the recovery of function among the heavy, former users do not appear to have clinical underpinnings
the absence of significant cognitive differences between the former users and the control group suggests that residual marihuana-associated deficits observed among current users are transient phenomena and that regular, heavy use does not produce irreversible, neurocognitive effects
Frohlich J, Van Horn J. 2014. “Reviewing the ketamine model for schizophrenia” J Psychopharmacol 28: 287
http://www.ncbi.nlm.nih.gov/pubmed/24257811 in dropbox
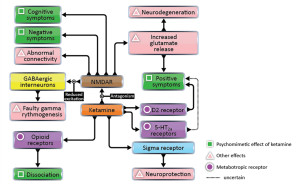
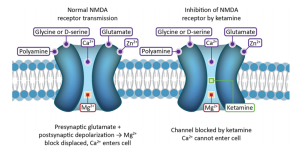
striatal regions suffer from hyperdopaminergia, while prefrontal cortical regions suffer from hypodopaminergia (Howes and Kapur, 2009; Lau et al., 2013)
Schizophrenic hypofrontality is hypothesized to lessen tonic dopamine release, and phasic dopamine release is in turn increased as a homeostatic compensation mechanism (Grace, 1991)
single photon emission computed tomography (SPECT) imaging of healthy subjects shows a strong ( r =0.96) correlation between negative symptoms and ketamine-induced changes in the binding of a [123I]CNS-1261 radiotracer to NMDARs (Stone et al., 2008)
dysfunctional role of cortico-limbocortico-thalamic circuitry in schizophrenia (Tsai and Coyle, 2002)
NMDAR is rarely considered a glycine receptor because it is glutamate that is released from the presynaptic terminal, with micromolar background levels of glycine sufficing for channel activation
Although findings have been mixed, many studies report a positive correlation between levels of dynorphins, endogenous kappa receptor ligands, in the cerebrospinal fluid of schizophrenia patients and psychotic symptoms (Tejeda et al., 2012)
ketamine and other uncompetitive NMDAR antagonists have a net positive effect on excitatory transmission by inducing excessive release of glutamate (Kim et al., 2011; Rowland et al., 2005; Stone et al., 2012) and acetylcholine (Giovannini et al., 1994; Hasegawa et al., 1993)
reduced levels of a protein linked to NMDAR function and working memory (Karlsgodt et al., 2011), dystrobrevin-binding-protein-1 (dysbindin) http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4204919/
the gene encoding dysbindin, and GRIN2B (Li and He, 2007a), the gene encoding the NR2B subunit of NMDAR, have been associated with schizophrenia
The growth factor neuregulin-1 (NRG1), its receptor, erbB4, and the scaffold protein known as postsynaptic density protein of 95 kDa (PSD-95); NRG1 inhibits prefrontal cortical NMDAR in humans, and schizophrenics have increased interactions between erbB4 and PSD-95 (Hahn et al., 2006)
D-amino acid oxidase (DAAO) is an enzyme that degrades D-serine by oxidation. Several single nucleotide polymorphisms (SNPs) in DAAO have been linked to schizophrenia (Chumakov et al., 2002). Additionally, G72 (also known as D-amino acid oxidase activator: DAOA) and G30, overlapping genes that interact with DAAO, have SNPs associated with schizophrenia (Li and He, 2007b: 30; Shinkai et al., 2007)
supplementing antipsychotic medication with D-serine alleviates the negative (p=0.02; pooled effect size (ES)=0.48), cognitive (p=0.007; ES=0.42), and total ( p=0.02; ES=0.4) symptoms of schizophrenia (Tsai and Lin, 2010)
significant reductions in all psychiatric groups (schizophrenic, depressive, and bipolar) for GRIN1 mRNA encoding the NR1 subunit of NMDAR in the dentate gyrus. Unique to the schizophrenia cohort was the left-lateralization of this finding (Law and Deakin, 2001)
enhanced sensitivity to the dopaminergic effects of amphetamine challenge is also characteristic of schizophrenia patients, as revealed by PET and SPECT imaging (Breier et al., 1997; Laruelle et al., 1996)
increased glutamate release, induced by ketamine, explains observed frontal deactivation (Deakin et al., 2008)
a PET study of schizophrenia patients reported fewer functional interactions between brain regions than controls, including reduced correlations between activation of anterior and posterior regions and between thalamic and cortical regions (Volkow et al., 1988)
Anatomical networks in patients are more sparsely connected when analyzed both in terms of axonal connectivity and cortical thickness (an indirect measure of the former) (Bassett et al., 2008; Wang et al., 201 1; Zalesky et al., 201 1). Resting-state
functional networks in patients show reduced connection density (“clustering”), fewer “hub” regions with very high interconnectivity, and reduced efficiency (“small-worldness”) of connections (Lynall et al., 2010)
(DMN), a set of brain regions deactivated by cognitive tasks and forming a functional module in resting state functional networks observed with fMRI (Sporns, 2012), has increased connectivity in schizophrenia patients. What is more, the extent of
strengthened connectivity within the DMN is correlated with the severity of psychiatric symptoms (Whitfield-Gabrieli et al., 2009)
TMS-evoked waves of excitation propagate longer and farther in schizophrenia patients than controls (Frantseva et al., 2012)
Postmortem studies of PFC tissue from schizophrenic patients reinforce the above findings by revealing reduced numbers of GABAer gic interneurons expressing GAD67 (Hashimoto et al., 2003; Volk et al., 2000), parvalbumin (Hashimoto et al., 2003), and GABA transporter-1 (GAT-1) mRNA (Volk et al., 2001). PFC chandelier cells, GABAergic neurons that regulate pyramidal cell output, have lower axon cartridge (terminal array) density in schizophrenia patients (Pierri et al., 1999)
several lines of evidence converge to implicate the thalamic reticular nucleus in the pathology of schizophrenia (Ferrarelli and Tononi, 2011)
increased global based connectivity of six clusters are associated with positive symptoms, whereas constant or decreased GBC in the dorsal and medial striatum and thalamus is associated with negative symptoms (Driesen et al., 2013)
while gamma oscillations in schizophrenia are often reduced, increases have also been observed (Bucci et al., 2007). Like resting-state GBC, decreases are often associated with negative symptoms and increases are often associated with positive symptoms (Figure 3) (Baldeweg et al., 1998; Bucci et al., 2007; Kehrer et al., 2008; Spencer et al., 2008: 2)
cognitive control induced modulation of prefrontal gamma band activity, correlated with cognitive control demand and task performance in healthy controls, is not observed in schizophrenia patients (Cho et al., 2006)
N-back tests of working memory induce excessive frontal gamma band activity in schizophrenia patients, whose performance is worse than that of healthy controls (Barr et al., 2010, 2011). One might draw a crude analogy between the schizophrenic PFC and an overworked engine: frontal networks in schizophrenia appear inflexible and inefficient, thus serving as weaker substrates for working memory
patients have higher absolute gamma synchrony relative to controls (Williams et al., 2009) Cortical networks in schizophrenia appear rigid and inflexible, as judged from their gamma band activity
a heartbeat that is too regular is linked to cardiac pathologies such as congestive heart failure (Ho et al., 2011; Norris et al., 2008; Poon and Merrill, 1997)
schizophrenia patients’ oscillations in lower frequencies bands include both abnormal increases and decreases in power during both induced and resting oscillations (Uhlhaas and Singer, 2011)
NMDAR antagonists that are nonspecific or specific to the NR2A subunit increase gamma power in rat cortex, whereas antagonists specific to other NMDAR subunits have weak effects (Kocsis, 2012)
the general theta-to-gamma shift observed in schizophrenia is replicated using ketamine (Ehrlichman et al., 2009). Ketamine also increases the power of 20–29 Hz oscillations in the beta band in the association cortex and PFC, a finding likely compatible with increased frontal and parietal resting beta oscillations in schizophrenia patients with auditory hallucinations (Lee et al., 2006; Mulert et al., 2011; Roopun et al., 2008)
Fu Q, Heath A, ZBucholz K, Nelson E, Goldberg J, Lyons M, True W, Jacob T, Tsuang M, Eisen S. 2002. “Shared genetic risk of major depression, alcohol dependence, and marijuana dependence: contribution of antisocial personality disorder in men” Arch Gen Psychiatry 59(12): 1125-32
http://www.ncbi.nlm.nih.gov/pubmed/12470129/
Genetic effects on ASPD accounted for 38%, 50%, and 58% of the total genetic variance in risk for MD, AD, and MJD, respectively … after controlling for ASPD, genetic correlations of MD with AD and with MJD not statistically significant
Fu C, Williams S, Brammer M, Suckling J, Kim J, Cleare A, Walsh N, Mitterschiffthaler M, Andrew C, Pich E, Bullmore E. 2007. “Neural responses to happy facial expressions in major depression following antidepressant treatment” Am J Psychiatry 164(4): 599-607
http://www.ncbi.nlm.nih.gov/pubmed/17403973?dopt=Abstract
Impairments in the neural processing of happy facial expressions in depression were evident in the core regions of affective facial processing, which were reversed following treatment. These data complement the neural effects observed with negative affective stimuli
Fusar-Poli P, Crippa JA, Bhattacharyya S, Borgwardt SJ, Allen P, Martin-Santos R, Seal M, Surguladze SA, O’Carrol C, Atakan Z, Zuardi AW, McGuire PK. 2009. “Distinct effects of {delta}9-tetrahydrocannabinol and cannabidiol on neural activation during emotional processesing.” Arch Gen Psychiatry 66(1):95-105
http://www.ncbi.nlm.nih.gov/pubmed/19124693 in dropbox
“Delta9-Tetrahydrocannabinol increased anxiety, as well as levels of intoxication, sedation, and psychotic symptoms, whereas there was a trend for a reduction in anxiety following administration of CBD”
Fusar-Poli P, Allen P, Bhattacharyya S, Crippa JA, Mechelli A, Borgwardt S, Martin-Santos R, Seal ML, O’Carrol C, Atakan Z, Zuardi AW, McGuire P. 2010. “Modulation of effective connectivity during emotional processing by Delta 9-tetrahydrocannabinol and cannabidiol” International Journal of Neuropsychopharmacology 13(4):421-32
http://www.ncbi.nlm.nih.gov/pubmed/19775500
“BMS identified a model with driving inputs entering via the anterior cingulate and forward intrinsic connectivity between the amygdala and the anterior cingulate as the best fit. CBD but not Delta 9-THC disrupted forward connectivity between these regions during the neural response to fearful faces”
Fuster D, Cheng D, Allensworth-Davies D, Palfai T, Samet J, Saitz R. 2014. “No detectable association between frequency of marijuana use and health or healthcare utilization among primary care patients who screen positive for drug use” J Gen Intern Med 29(1): 133-9
http://www.ncbi.nlm.nih.gov/pubmed/24048656
All 589 participants reported recent drug use: marijuana 84 % (29 % daily, 55 % less than daily), cocaine 25 %, opioid 23 %, other drugs 8 %; 58 % reported exclusive marijuana use. Frequency of marijuana use was not significantly associated with emergency department use {adjusted odds ratio [AOR] 0.67, [95 % confidence interval (CI) 0.36, 1.24] for daily; AOR 0.69 [95 % CI 0.40,1.18] for less than daily versus no use}, hospitalization [AOR 0.79 (95 % CI 0.35, 1.81) for daily; AOR 1.23 (95 % CI 0.63, 2.40) for less than daily versus no use], any comorbidity [AOR 0.62, (95 % CI 0.33, 1.18) for daily; AOR 0.67 (95 % CI 0.38, 1.17) for less than daily versus no use] or health status (adjusted mean EuroQol 69.1, 67.8 and 68.0 for daily, less than daily and none, respectively, global p = 0.78).
Gaffal E, Cron M, Glodde N, Tuting T. 2013. “Anti-inflammatory activity of topical THC in DNFB-mediated mouse allergic contact dermatitis independent of CB1 and CB2 receptors” Allergy 68(8): 994-1000
http://www.ncbi.nlm.nih.gov/pubmed/23889474/
Topical THC application effectively decreased contact allergic ear swelling and myeloid immune cell infiltration not only in wild-type but also in CB1/2 receptor-deficient mice. We found that THC (1) inhibited the production of IFNγ by T cells, (2) decreased the production of CCL2 and of IFNγ-induced CCL8 and CXL10 by epidermal keratinocytes and (3) thereby limited the recruitment of myeloid immune cells in vitro in a CB1/2 receptor-independent manner
Topically applied THC can effectively attenuate contact allergic inflammation by decreasing keratinocyte-derived pro-inflammatory mediators that orchestrate myeloid immune cell infiltration
Galea S, Ahern J, Resnick H, Kilpatrick D, Bucuvalas M, Gold J, Vlahov D. 2002. “Psychological sequelae of the September 11 terrorist attacks in New York City” N Engl J Med 346(13): 982-7
http://www.ncbi.nlm.nih.gov/pubmed/11919308/
Among 1008 adults interviewed, 7.5 percent reported symptoms consistent with a diagnosis of current PTSD related to the attacks, and 9.7 percent reported symptoms consistent with current depression (with “current” defined as occurring within the previous 30 days). Among respondents who lived south of Canal Street (i.e., near the World Trade Center), the prevalence of PTSD was 20.0 percent. Predictors of PTSD in a multivariate model were Hispanic ethnicity, two or more prior stressors, a panic attack during or shortly after the events, residence south of Canal Street, and loss of possessions due to the events. Predictors of depression were Hispanic ethnicity, two or more prior stressors, a panic attack, a low level of social support, the death of a friend or relative during the attacks, and loss of a job due to the attacks.
Galve-Roperh I, Aguado T, Palazuelos J, Guzman M. 2008. “Mechanisms of control of neuron survival by the endocannabinoid system” Curr Pharm Des 14(23): 2279-88
http://www.ncbi.nlm.nih.gov/pubmed/18781978
Thus, CB(1) receptor activation has been shown to protect neurons from acute brain injury as well as in neuroinflammatory conditions and neurodegenerative diseases
Cannabinoid neuroprotective activity relies on the inhibition of glutamatergic neurotransmission and on other various mechanisms, and is supported by the observation that the brain overproduces endocannabinoids upon damage. Coupling of neuronal CB(1) receptors to cell survival routes such as the phosphatidylinositol 3-kinase/Akt and extracellular signal-regulated kinase pathways may contribute to cannabinoid neuroprotective action. These pro-survival signals occur, at least in part, by the cross-talk between CB(1) receptors and growth factor tyrosine kinase receptors. Besides promoting neuroprotection, a role for the endocannabinoid system in the control of neurogenesis from neural progenitors has been put forward. In addition, activation of CB(2) cannabinoid receptors on glial cells may also participate in neuroprotection by limiting the extent of neuroinflammation
Galynker I, Cai J, Ongseng F, Finestone H, Dutta E, Serseni D. 1998. “Hypofrontality and negative symptoms in major depressive disorder” J Nucl Med 39(4):608-12
http://www.ncbi.nlm.nih.gov/pubmed/9544664
Major depressive disorder (MDD) group had significantly lower rCBF in the frontal cortex and cinglulate gyrus due to differences in dorsolateral prefrontal cortex bilaterally (right F = 7.69, p = 0.01; left F = 8.41, p = 0.01) in the right orbitofrontal cortex (F = 6.79, p = 0.02) and in the cingulate gyrus (F = 5.34, p = 0.03). The MDD group also had lower rCBF in the posterior cortical structures (MANOVA, p = 0.072) due to decreased perfusion in the right parietal cortex (F = 7.54, p = 0.01). negative correlations between the SANS total score and rCBF in both the left dorsolateral prefrontal cortex (Pearson’s correlation coefficient r = .-67, p < 0.05) and the left anterior temporal cortex (r = -0.71, p < 0.01) in MDD patients. Additionally, there were positive correlations between HRSD scores and rCBF in the left anterior temporal (r = 0.71, p < 0.01), left dorsolateral prefrontal (r = 0.70, p < 0.01), right frontal (r = 0.82, p < 0.01) and right posterior temporal (r = 0.74, p < 0.01) cortices. Hypofrontality/decreased perfusion correlates with negative symptoms
Ganon-Elazar E, Akirav I. 2012. “Cannabinoids prevent the development of behavioral and endocrine alterations in a rat model of intense stress” Neuropsychopharmacology 37(2): 456-466
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3242307/
WIN (0.5mg/kg) administered intraperitoneally 2 or 24h (but not 48h) after SPS prevented the trauma-induced alterations in IA conditioning and extinction, ASR potentiation, and HPA axis inhibition
(i) there may be an optimal time window for intervention treatment with cannabinoids after exposure to a highly stressful event, (ii) some of the preventive effects induced by WIN are mediated by an activation of CB1 receptors in the BLA, and (iii) cannabinoids could serve as a pharmacological treatment of stress- and trauma-related disorders.
Ganzel B, Casey B, Glover G, Voss H, Temple E. 2007. “The Aftermath of 9/11: Effect of Intensity and Recency of Trauma on Outcome” Emotion 7(2): 227-238
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2759706/
More than three years after the terrorist attacks, bilateral amygdala activity in response to viewing fearful faces compared to calm ones was higher in people who were within 1.5 miles of the World Trade Center on 9/11, relative to those who were living more than 200 miles away
This activity mediated the relationship between group status and current symptoms of posttraumatic stress disorder. In turn, the effect of group status on both amygdala activation (fearful vs. calm faces) and current symptoms was statistically explained by time since worst trauma in lifetime and intensity of worst trauma, as indicated by reported symptoms at time of the trauma. These data are consistent with a model of heightened amygdala reactivity following high-intensity trauma exposure, with relatively slow recovery.
increased activity in the amygdala is the most consistently observed neural correlate of PTSD as probed with emotional faces, with some evidence that this activation is localized to subregions of the amygdala and is associated with alterations in medial prefrontal activity.
increased amygdala activation predicted increased current symptoms
Garcia-Bueno B, caso J, Leza J. 2008. “Stress as a neuroinflammatory condition in brain: damaging and protective mechanisms” Neurosci Biobehav Rev 32(6): 1136-51
http://www.ncbi.nlm.nih.gov/pubmed/18468686/
Several neuropsychiatric diseases are related with stress (posttraumatic stress disorder, major depressive disorder, anxiety disorders, schizophrenia) and stress exposure modifies the onset and evolution of some neurological diseases (neurodegenerative diseases). It is accepted that brain inflammatory responses contribute to cell damage during these illnesses
pro-inflammatory response in the brain and other systems mainly characterized by a complex release of several inflammatory mediators such as cytokines, prostanoids, free radicals and transcription factors
Genetic Risk and Outcome in Psychosis (GROUP) Investigators. 2011. “Evidence that familial liability for psychosis is expressed as differential sensitivity to cannabis an analysis of patient-sibling and sibling-control pairs” Arch Gen Psychiatry 68(2): 138-147
http://archpsyc.jamanetwork.com/article.aspx?articleid=211037
a sibling-based design may aid in teasing apart moderation (genetic control of sensitivity to the psychotomimetic effects of cannabis) from mediation (genetic control of exposure to cannabis)
Both patients and siblings displayed higher rates of cannabis use, suggesting that some of the apparent differential sensitivity to the psychotomimetic effects of cannabis may in fact represent a mechanism of psychotic disorder liability contributing not only to sensitivity for cannabis but also to the tendency to start using cannabis (gene-environment correlation).50 However, the results of the cross-sibling, cross-trait analyses examining whether cannabis use and psychosis liability were associated with each other across siblings did not support a gene-environment correlation. This is in agreement with a previous study examining this issue51 and with genetic epidemiology findings indicating that cannabis use in (early) adolescence—the relevant timing of exposure with regard to psychotic disorder52—is influenced by environmental rather than genetic factors.53 Thus, it is likely that the findings reflect, at least in part, moderation of the psychotomimetic effects of cannabis by familial liability to psychotic disorder.
Similarly, cannabis use assessed by urinalysis increased the sibling correlation by an approximate factor of 10 for positive symptoms and a factor of 2 (not statistically significant) for negative symptoms (Table 5).
An important issue revealed by this study is that while the relative effect sizes of differential sensitivity were high, absolute effect sizes, for example, of cannabis on schizotypy in unaffected siblings, were small
differential sensitivity to cannabis was demonstrated in the form of greater response to recent cannabis use in individuals at higher-than-average risk for psychotic disorder and related disorders
George K, Saltman L, Stein G, Lian J, Zurier R. 2008. “Ajulemic acid, a nonpsychoactive cannabinoid acid, suppresses osteoclastogenesis in mononuclear precursor cells and induces apoptosis in mature osteoclast-like cells” J Cell Physiol 214(3): 714-20
http://www.ncbi.nlm.nih.gov/pubmed/17786950/
Oral administration of ajulemic acid (AjA), a cannabinoid acid devoid of psychoactivity, prevents joint tissue injury in rats with adjuvant induced arthritis
Reduction by AjA of both monocyte growth and osteoclast formation was associated with apoptosis, assayed by annexin V and propidium iodide staining, and caspase activity. The anti-osteoclastogenic effects of AjA did not require the continuous presence of AjA in the cell cultures. Based on these findings, we propose that AjA or other nonpsychoactive synthetic analogs of Cannabis constituents may be useful therapy for diseases such as RA and osteoporosis in which bone resorption is a central feature.
Gerritsen L, Velzen L, Schmaal L, van der Graaf Y, van der Wee N, van Tol M-J, Penninx B, Geerlings M. 2015. “Childhood maltreatment modifies the relationship of depression with hippocampal volume” Psychological Medicine 45(16): 3517-3526
http://journals.cambridge.org/action/displayAbstract?fromPage=online&aid=10017663&fulltextType=RA&fileId=S0033291715001415
262 participants from the Netherlands Study of Depression and Anxiety (NESDA) (mean age 37 years, 32% male) and 636 participants from the SMART-Medea study (mean age 61 years, 81% male)
In both cohorts CM was not significantly associated with hippocampal volume. After pooling the data MDD was associated with smaller hippocampal volume (B = −138.90 mm3, p = 0.05) and the interaction between MDD and CM reached significance (p = 0.04); in participants with CM, MDD was related to smaller hippocampal volume (NESDA: B = −316.8 mm3, p = 0.02; SMART: B = −407.6, p = 0.046), but not in participants without CM (p > 0.05)
Gertsch J, Leonti M, Raduner S, Racz I, Chen J-Z, Xie X-Q, Altmann K-H, Karsak M, Zimmer A. 2008 “Beta-caryophyllene is a dietary cannabinoid” Proc Natl Acad Sci U S A 105(26): 9099-9104
http://www.pnas.org/content/105/26/9099
Although the CB1 receptor is responsible for the psychomodulatory effects, activation of the CB2receptor is a potential therapeutic strategy for the treatment of inflammation, pain, atherosclerosis, and osteoporosis. Here, we report that the widespread plant volatile (E)-β-caryophyllene [(E)-BCP] selectively binds to the CB2 receptor (K i = 155 ± 4 nM) and that it is a functional CB2 agonist
common constituent of the essential oils of numerous spice and food plants and a major component inCannabis
Upon binding to the CB2receptor, (E)-BCP inhibits adenylate cylcase, leads to intracellular calcium transients and weakly activates the mitogen-activated kinases Erk1/2 and p38 in primary human monocytes. (E)-BCP (500 nM) inhibits lipopolysaccharide (LPS)-induced proinflammatory cytokine expression in peripheral blood and attenuates LPS-stimulated Erk1/2 and JNK1/2 phosphorylation in monocytes. Furthermore, peroral (E)-BCP at 5 mg/kg strongly reduces the carrageenan-induced inflammatory response in wild-type mice but not in mice lacking CB2 receptors, providing evidence that this natural product exerts cannabimimetic effects in vivo. These results identify (E)-BCP as a functional nonpsychoactive CB2 receptor ligand in foodstuff and as a macrocyclic antiinflammatory cannabinoid in Cannabis
Giaconia R, Reinherz H, Hauf A, Paradis A, Wasserman M, Langhammer D. 2000. “Comorbidity of substance use and post-traumatic stress disorders in a community sample of adolescents” Am J Orthopsychiatry 70(2): 253-62
http://www.ncbi.nlm.nih.gov/pubmed/10826037
A study of 384 18-year-old adolescents living in the community demonstrated a frequent co-occurrence of substance use disorders and post-traumatic stress disorder. Multiple pathways appeared to lead to this comorbidity, which was associated with widespread psychological impairment that might have serious developmental consequences
Giacoppo S, Galuppo M, Pollastro F, Grassi G, Bramanti P, Mazzon E. 2015. “A new formulation of cannabidiol in cream shows therapeutic effects in a mouse model of experimental autoimmune encephalomyelitis” Daru 23: 48
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4618347/
topical 1 % CBD-cream may exert neuroprotective effects against EAE, diminishing clinical disease score (mean of 5.0 in EAE mice vs 1.5 in EAE + CBD-cream), by recovering of paralysis of hind limbs and by ameliorating histological score typical of disease (lymphocytic infiltration and demyelination) in spinal cord tissues. Also, 1 % CBD-cream is able to counteract the EAE-induced damage reducing release of CD4 and CD8α T cells (spleen tissue localization was quantified about 10,69 % and 35,96 % of positive staining respectively in EAE mice) and expression of the main pro-inflammatory cytokines as well as several other direct or indirect markers of inflammation (p-selectin, IL-10, GFAP, Foxp3, TGF-β, IFN-γ), oxidative injury (Nitrotyrosine, iNOS, PARP) and apoptosis (Cleaved caspase 3)
It is known that when orally given, due to a marked first-pass effect, CBD bioavailability ranges between values of 13 and 19 % [37], hence the oral route is not ideal for the therapeutic delivery of CBD
1 % CBD-cream treatment acts counteracting leukocyte infiltration and microglial activation and improving neuroinflammation status. Hystopathology of spinal cord samples supported this view showing a marked remyelination following 1 % CBD-cream treatment
As expected we found increased levels of proinflammatory mediators including IL-1β, TNF-α, IL-6, IFN-γ and TGF-β in spinal cord from EAE mice. Remarkably, 1 % CBD-cream treatment significantly attenuates the expression of all these inflammatory markers and also regulates immune tolerance by increasing the production of antinflammatory cytokines, like IL-10. This confirms that CBD was able to decrease TNF-α, IL-2 and IFN-γ release from activated splenocytes and macrophages as reported in other studues [47–49]
it’s probably that all these mediators are produced by Th17 cells, that play a key role in in autoimmune neuroinflammation and EAE development [50]. Also, it is generally considered that Th17-mediated inflammation is characterized by neutrophil recruitment into the CNS and neurons killing. In this context, mainly IFN-γ seems to have a pathological role in the development of this autoimmune disease [51]. Our results show that the MOG-induced EAE causes significant increases in IFN-γ expression. 1 % CBD-cream administration, significantly diminished the expression of this cytokine
To verify if treatment with CBD can modulate the production of Treg cells, we evaluated the expression of the transcription factor Foxp3, as an indirect marker of Treg. In accordance with a previous study [52], we confirmed a clear engagement of Treg cells during EAE, while treatment with CBD-cream is able to deplete Foxp3 positive cells. This leads to think that the immune system response is somehowe restrained by topic application of 1 % CBD-cream in EAE affected mice in correlation with a lower degree of autoimmune cells activity in these animals. Also, it is possible that 1 % CBD-cream stimulates Th0 cell to develop into a Treg phenotype
we demonstrated that CBD reduces the expression of iNOS and nitrotyrosine in tissues from EAE treated mice, suggesting that this compound, at least in part, may be responsible for the reduction of cytokines production, that in turn counteract the rise of iNOS levels and the downstream cascade of events triggered by the inflammatory process, reducing thus oxidative stress
CBD has the capability to interfere with EAE-induced neuronal apoptotic death, attenuating, or even preventing the activation of cellular molecular mechanisms triggered by severe damage
Gil T, Calev A, Greenberg D, Kugelmass S, Lerer B. 1990. “Cognitive functioning in post-traumatic stress disorder” Journal of Traumatic Stress 3(1):29-45
http://onlinelibrary.wiley.com/doi/10.1002/jts.2490030104/abstract in dropbox
“intelligence of PTSD patients and psychiatric controls was average and had deteriorated significantly by the time of current testing. not secondary to alcohol, drug abuse, or head injury. The results suggest a cognitive impairment in post-traumatic patients.”
13 point drop from premorbid IQ
Gilbert G, Kim H, Waataja J, Thayer S. 2007. “Delta9-tetrahydrocannabinol protects hippocampal neurons from excitotoxicity” Brain Res 1128(1): 61-9
http://www.ncbi.nlm.nih.gov/pubmed/17140550/
Excitotoxic neuronal death underlies many neurodegenerative disorders. Because cannabinoid receptor agonists act presynaptically to inhibit glutamate release, we examined the effects of Win 55212-2, a full agonist at CB(1) receptors, and Delta(9)-tetrahydrocannabinol (THC), a partial agonist, on the survival of neurons exposed to an excitotoxic pattern of synaptic activity
Win 55212-2 (100 nM) and THC (100 nM) significantly reduced 0.1 mM [Mg(2+)](o)-induced cell death by 77 +/- 11% and 84 +/- 8%, respectively… the protection afforded by THC was not significantly different from that produced by Win 55212-2, suggesting that attenuation without a complete block of excitatory activity is sufficient for neuroprotection
When cultures were pretreated for 24 h with Win 55212-2 (100 nM) or THC (100 nM), inhibition of 0.1 mM [Mg(2+)](o)-induced toxicity was significantly reduced to 39 +/- 19% and 45 +/- 13%, respectively. Thus, desensitization of CB(1) receptors diminishes the neuroprotective effects of cannabinoids
Gilbertson M, Shenton M, Ciszewski A, Kasai K, Lasko N, Orr S, Pitman R. 2002. “Smaller hippocampal volume predicts pathologic vulnerability to psychological stress” Nature Neuroscience 5:1242-1247
http://www.nature.com/neuro/journal/v5/n11/full/nn958.html
Disorder severity in PTSD patients who were exposed to trauma was negatively correlated with the hippocampal volume of both the patients and the patients’ trauma-unexposed identical co-twin
Gill J, Luckenbaugh D, Charney D, Vythilingam M. 2010. “Sustained elevation of serum interleukin-6 and relative insensitivity to hydrocortisone differentiates posttraumatic stress disorder with and without depression” Biol Psychiatry 68(11): 999-1006
http://www.ncbi.nlm.nih.gov/pubmed/20951370
Only subjects with PTSD + MDD exhibited higher, overnight serum IL-6 levels compared with individuals with PTSD – MDD (p < .01) and healthy control subjects (p < .001). Peak overnight IL-6 levels positively correlated with severity of PTSD (r = .56, p < .01) and depressive symptoms (r = .54, p < .01)
Giuffrida A, Parsons L, Kerr T, Fonseca F, Navarro M, Piomelli D. 1999. “Dopamine activation of endogenous cannabinoid signaling in dorsal striatum” Nat Neurosci 2(4): 358-63
http://www.ncbi.nlm.nih.gov/pubmed/10204543
Neural activity stimulated the release of anandamide, but not of other endogenous cannabinoids such as 2-arachidonylglycerol. Moreover, anandamide release was increased eightfold over baseline after local administration of the D2-like (D2, D3, D4) dopamine receptor agonist quinpirole, a response that was prevented by the D2-like receptor antagonist raclopride
The endocannabinoid system therefore may act as an inhibitory feedback mechanism countering dopamine-induced facilitation of motor activity
Giuffrida A, Leweke F, Gerth C, Schreiber D, Koethe D, Faulhaber J, Klosterkotter J, Piomelli D. 2004. “Cerebrospinal anandamide levels are elevated in acute schizophrenia and are inversely correlated with psychotic symptoms” Neuropsychopharmacology 29: 2108-2114
http://www.ncbi.nlm.nih.gov/pubmed/15354183 in dropbox
CSF anandamide levels are eight-fold higher in antipsychotic-naive first-episode paranoid schizophrenics (n-47) than healthy controls (n=84), dementia patients (n=13) or affective disorder patients (n=22) … absent in schizophrenics treated with typical antipsychotics (n=37) but not in those treated with atypical antipsychotics (n=34)
in nonmedicated acute schizophrenics, CSF anandamide is negatively correlated with psychotic symptoms. These results suggest that anandamide elevation in acute paranoid schizophrenia may reflect a compensatory adaptation to the disease state
In contrast to the elevation observed with anandamide, CSF levels of OEA were identical in antipsychotic-naı ¨ve schizophrenics and control subjects (controls, 1.011 7 0.136 pmol/ml, n ¼ 81; antipsychotic-naı ¨ve schizophrenics, 1.11670.217 pmol/ml, n ¼ 37,P ¼ 0.624, Kruskal–Wallis test), while CSF levels of PEA were lower in antipsychotic-naı¨ve
Previous studies have suggested that anandamide release in the brain may serve as an inhibitory feedback signal countering dopamine activation of motor behavior (Giuf-frida et al , 1999; Beltramo et al , 2000). We hypothesized therefore that anandamide might also serve as a homeo-static function in acute schizophrenia, in which exaggerated dopaminergic transmission has been documented (Abi-Dargham et al , 2000; Meyer-Lindenberg et al , 2002)
CSF anandamide levels were negatively correlated with PANSS scores (PANSS-T: cumulative value for all symptom complexes; PANSS-P: positive symptoms; PANSS-N: negative symptoms; and PANSS-G: general symptoms) (Figure 2) (PANSS-T: rS ¼0.452, P ¼ 0.001; PANSS-P:r S ¼0.347; P ¼ 0.017; PANSS-N: r S ¼0.409, P ¼ 0.004; PANSS-G: r S ¼0.426, P ¼ 0.003; n ¼ 47)
history of cannabis use in the group of healthy controls and antipsychotic-naı¨ve schizophrenics was comparable to some extent and no overall effect of cannabis use on the primary end point of this study was observed
the elevation in anandamide concentration was unlikely to be caused by a generalized alteration in lipid signaling, which is hypothesized to occur in schizophrenia (Horrobin 1998), because the levels of two closely related analogs of anandamide, OEA and PEA, were not increased in schizophrenics
D2-like receptors initiates anandamide synthesis in limbic and motor areas of the rat brain (Giuffrida et al , 1999; Ferrer et al, 2003)
Phar-macological experiments in the rat have suggested that anandamide release may serve as an inhibitory feedback signal countering dopamine activation of motor behavior (Giuffrida et al , 1999; Beltramo et al , 2000). The negative correlation of CSF anandamide with psychopathological symptoms in acute, antipsychotic-naı¨ve paranoid schizo-phrenics suggests that anandamide might play a similar adaptive role in acute schizophrenia
it is important to point out results from a Phase III clinical trial, which suggests that the CB1 antagonist rimonabant (SR141716A) has no therapeutic effect in schizophrenia and schizoaffec-tive disorders (Arvanitis et al , 2002)
Glass C, Saijo K, Winner B, Marchetto M, Gage F. 2010. “Mechanisms underlying inflammation in neurodegeneration” Cell 140(6): 918-934
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2873093/
Although inducers of inflammation may be generated in a disease-specific manner, there is evidence for a remarkable convergence in the mechanisms responsible for the sensing, transduction, and amplification of inflammatory processes that result in the production of neurotoxic mediators
Sustained inflammation resulting in tissue pathology implies persistence of an inflammatory stimulus or a failure in normal resolution mechanisms
Although some inflammatory stimuli induce beneficial effects (e.g., phagocytosis of debris and apoptotic cells), and inflammation is linked to tissue repair processes, uncontrolled inflammation may result in production of neurotoxic factors that amplify underlying disease states
Roles of TLR2 and TLR4 have recently been established in the pathogenesis of several chronic inflammatory diseases in animal models, and specific TLR4 polymorphisms are associated with several human age-related diseases, including atherosclerosis, type 2 diabetes, and rheumatoid arthritis, raising the question of whether these receptors also contribute to inflammatory programs associated with neurodegenerative disease (Balistreri et al., 2009) (TLR= toll-like receptor)
purinergic receptors are expressed on microglia and astrocytes and are capable of responding to ATP released from cells following cell death, traumatic injury, or ischemia (Di Virgilio et al., 2009). Microglia and astrocytes also express a number of so-called “scavenger receptors” that have been demonstrated to be involved in the uptake of a number of substrates, including oxidized proteins, lipids, and apoptotic cells, and that may also contribute to cell signaling (Husemann et al., 2002).
TLRs couple to signaling adaptor systems that are defined by the MyD88 and TRIF signal adaptor proteins, resulting in activation of downstream kinases including IkB kinases and MAP kinases. These in turn control the activities of multiple, signal-dependent transcription factors that include members of the NF-κB, AP-1, and interferon regulator factor (IRF) families. These factors work in a combinatorial manner to regulate hundreds of genes, depending on the target cell that is activated
To bring about an effective immune response, the initial detection of a microbial pathogen must be amplified to recruit additional cells to sites of infection, induce antimicrobial activities, and initiate the development of adaptive immunity. Important subsets of highly induced genes thus include cytokines (e.g., TNF-α, IL-1β) that are amplifiers of the program of inflammation and chemokines (e.g., MCP-1) that serve to recruit additional immune cells. In addition, genes that encode proteins with antimicrobial activities (e.g., iNOS) are induced, as are genes that influence substrate metabolism, protein synthesis, cell motility, phagocytosis, intracellular killing, and antigen presentation. The generation of reactive oxygen species (ROS), e.g., through the NADPH oxidase system, is an important antimicrobial mechanism but also exemplifies a system that can result in collateral damage to tissue
Microglia appear to be the major initial sensors of danger or stranger signals recognized by TLR4, and they secrete inflammatory mediators such as TNF-α and IL-1β that can act on astrocytes to induce secondary inflammatory responses (Saijo et al., 2009). Microglia are thus likely to play critical roles in establishing and maintaining inflammatory responses in the context of neurodegenerative diseases.
The distinct pathways for production of inducers of inflammation—such as Aβ, α-synuclein, mutant SOD1, and myelin peptide mimetic—and the specific anatomical locations at which these processes occur are likely determinants of the specific pathologicial features of each disease. Remarkably, however, once inducers are generated, there appears to be considerable convergence in the sensor, transducer, and effector mechanisms that lead to amplification of inflammatory responses, neurotoxicity, and neuronal death. Activation of innate immune cells in the CNS, such as microglia and astrocytes, is one of the universal components of neuroinflammation (–). In particular, TLRs and other pattern recognition receptors expressed on microglia are likely to play significant roles in initiating inflammatory responses that are further amplified by astrocytes. Similarly, signal transduction pathways downstream of these receptors that regulate the activities of the transcription factors NF-κB and AP-1 appear to play general roles in mediating the production of amplifiers and effector molecules, such as cytokines (e.g., TNF-α, IL-1β, and IL-6), ROS, and NO. Several of these factors could be general neurotoxic factors for all of the neurodegenerative diseases discussed above. However, the marked involvement of the adaptive immune system clearly distinguishes MS from AD, PD, and ALS, although T cells do seem to play a neuroprotective role in ALS
Although inhibition of neuroinflammation may not alter the underlying cause of disease, it may reduce the production of factors that contribute to neurotoxicity, thereby resulting in clinical benefit
Glocker E, Kotlarz D, Klein C, Shah N, Grimbacher B. 2011. “IL-10 and IL-10 receptor defects in humans” Ann N Y Acad Sci 1246: 102-7
http://www.ncbi.nlm.nih.gov/pubmed/22236434
Inflammatory bowel disease (IBD), which includes Crohn’s disease (CD) and ulcerative colitis (UC), is chronic in nature and is characterized by abdominal pain, diarrhea, bleeding, and malabsorption. It is considered a complex multigenic and multifactorial disorder that results from disturbed interactions between the immune system and commensal bacteria of the gut. Recent work has demonstrated that IBD with an early-onset within the first months of life can be monogenic: mutations in IL-10 or its receptor lead to a loss of IL-10 function and cause severe intractable enterocolitis in infants and small children. Both IL-10 and IL-10 receptor deficiency can be successfully treated by hematopoietic stem cell transplantation
Golub Y, Kaltwasser S, Mauch C, Herrmann L, Schmidt U, Holsboer F, Czisch M, Wotjak C. 2011. “Reduced hippocampus volume in the mouse model of Posttraumatic Stress Disorder” Journal of Psychiatric Research 45(5): 650-659
http://www.journalofpsychiatricresearch.com/article/S0022-3956(10)00309-2/abstract
Exposure to a brief inescapable foot shock led to a volume reduction in both left HPC and right central amygdala two months later. This volume loss was mirrored by a down-regulation of growth-associated protein-43 (GAP43) in the HPC. Enriched housing decreased the intensity of trauma-associated contextual fear, independently of whether it was provided before or after the shock. Beyond that, enriched housing led to an increase in intracranial volume, including the lateral ventricles and the hippocampus, and to an up-regulation of GAP43 as revealed by MEMRI and Western blot analysis, thus partially compensating for trauma-related HPC volume loss and down-regulation of GAP43 expression
Gonzalez R, Swanson JM. 2012. “Long-term effects of adolescent-onset and persistent use of cannabis.” Proc Natl Acad Sci 109(40):15970-15971.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3479595/
past findings differ in two important ways from those reported by Meier et al.: they suggest that detrimental effects of cannabis may be specific to some neurocognitive functions (rather than general) and that cannabis-associated deficits may recover with abstinence (rather than persist)
nonregular users (50.6% of the total) showed no decline in IQ or neuropsychological performance — to whom do Meier’s findings apply?
de Graaf R, Radovanovic M, van Laar M, Fairman B, Degenhardt L, Aguilar-Gaxiola S, Bruffaerts R, Girolamo G, Fayyad J, Gureje O, Haro J, Huang Y, Kostychenko S, Lepine J-P, Matschinger H, Mora M, Neumark Y, Ormel J, Posada-Villa J, Stein D, Tachimori H, Wells J, Anthony J. 2010. “Early Cannabis Use and Estimated Risk of Later Onset of Depression Spells: Epidemiologic Evidence from the Population-based World Health Organization World Mental Health Survey Initiative” American Journal of Epidemiology
http://aje.oxfordjournals.org/content/early/2010/06/09/aje.kwq096.full
Association of before 17 cannabis with after 17 depression of 1.5. Controlling for childhood conduct problems reduces association to nonsignificance
The influences of family and parenting factors and childhood trauma were not studied; their confounding effects could be studied in future research
Grant I. 2003. “Non-acute (residual) neurocognitive effects of cannabis use: A meta-analytic study.” Journal of the International Neuropsychological Society 9(5):679-689
http://journals.cambridge.org/action/displayAbstract?fromPage=online&aid=154103
“alteration in learning and forgetting domains, lack of effect in other six… our results indicate that there might be decrements in the ability to learn and remember new information in chronic users, whereas other cognitive abilities are unaffected… small magnitude of these effect sizes suggests that they may have an acceptable margin of safety”
The results of our meta-analytic study failed to reveal a substantial, systematic effect of long-term, regular cannabis consumption on the neurocognitive functioning of users who were not acutely intoxicated. For six of the eight neuro-cognitive ability areas that were surveyed, the confidence intervals for the average effect sizes across studies over-lapped zero in each instance, indicating that the effect size could not be distinguished from zero
The two exceptions were in the domains of learning and forgetting. Here when we averaged across the 11 studies that had the most rigor-ous inclusion 0 exclusion criteria and the best designs, the effect size for learning was 2 .21 99%CI ( 2 .39, 2 .022) indicating a very small but discernible negative effect. This effect was slightly larger when all 15 studies were included [2 .24 99%CI (2 .41, 2 .064)]. Similarly, in the domain of forgetting (failure to recall or recognize) the average effect size was 2 .27 99%CI (2 .49, 2 .044), again suggesting a very small but measurable decrement
The “real life” impact of such a small and selective effect is questionable. In addition, it is important to note that most users across studies had histories of heavy long-term cannabis consumption. Therefore, these findings are not likely to generalize to more limited administration of cannabis compounds
another factor that may have contributed to the small ten-dency towards worse performance in the cannabis-using group might be attributable to what Pope et al. have called “residual effects.”
In nearly all instances, heavier can-nabis users were asked to abstain for a period of hours or days before testing. Therefore, many of them could have been at risk for “residual effects” or “abstinence phenom-ena,” which might have contributed to slight decrements in their performance. Given this likelihood, it is even more surprising that our meta-analytic study revealed so few effects
many studies had insufficient information about potential confounding fac-tors. These factors included recency of last cannabis expo-sure, extent of exposure to other drugs of abuse, presence of confounding neuropsychiatric factors (e.g., depression, anxiety, personality disorders, etc.), or other neuromedical risks that can independently affect brain function
It was surprising to find such few and small effects given that most of the potential biases inherent in our analy-ses actually increased the likelihood of finding a cannabis effect
Specifically, our use of a fixed effects model re-sulted in smaller confidence intervals for the effect sizes we computed, thus facilitating the discovery of statistically sig-nificant between-group differences. Moreover, many of the confounds inherent in the studies included in our analyses made it more likely for the cannabis using group to demon-strate poorer performance on neuropsychological tests than controls, irrespective of cannabis consumption. Finally, meta-analytic studies are generally criticized for including only investigations that have been published in peer-reviewed journals, because studies that report statistically significant findings are more likely to be published. This “file-drawer” bias can result in an underrepresentation of studies that did not find statistically significant results, therefore also in-creasing the likelihood of generating statistically significant effect sizes
Inspecting the effect sizes for neurocognitive domains by study, we noted that one study examining users in India (class-IV caste) of bhang and charas appeared to be a distant outlier
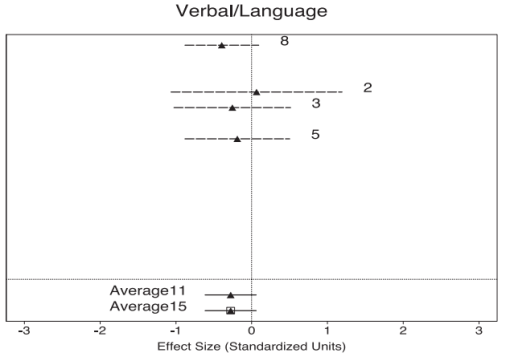
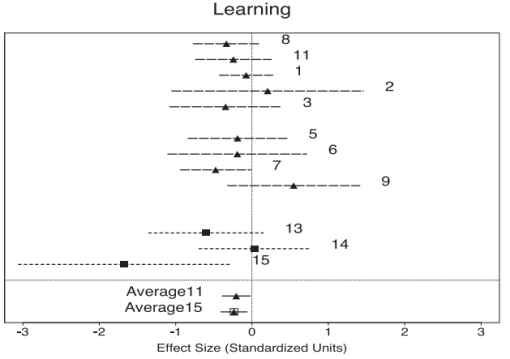
Grant B, Cho S, Goldstein R, Huang B, Stinson F, Saha T, Smith S, Dawson D, Pulay A, Pickering R, Ruan W. 2008. “Prevalence, correlates, disability and comorbidity of DSM-IV Borderline Personality Disorder: results from the wave 2 national epidemiologic survey on alcohol and related conditions” J Clin Psychiatry 69(4): 533-545
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2676679/
Prevalence of lifetime BPD was 5.9% (99% CI: 5.4–6.4). There were no differences in the rates of BPD among men (5.6%, 99% CI: 5.0–6.2) and women (6.2%, 99% CI: 5.6–6.9)
Prevalence of BPD in the general population is much greater than previously recognized, equal prevalent among men and women, and associated with considerable mental and physical disability, especially among women
Grant J, Lynskey M, Madden P, Nelson E, Few L, Bucholz K, Statham D, Martin N, Heath A, Agrawal A. 2015. “The role of conduct disorder in the relationship between alcohol, nicotine and cannabis use disorders” Psychological Medicine 45: 3505-3515
http://www.ncbi.nlm.nih.gov/pubmed/26281760 add to dropbox
Multivariate quantitative genetic analyses were conducted using data from 9577 individuals, including 3982 complete twin pairs and 1613 individuals whose co-twin was not interviewed (aged 24 – 37 years) from two Australian twin samples. Analyses examined the genetic correlation between alcohol dependence, nicotine dependence and cannabis abuse/dependence and the extent to which the correlations were attributable to genetic in fluences shared with conduct disorder
Additive genetic (a2 = 0.48– 0.65) and non-shared environmental factors explained variance in substance use dis-orders… ll substance use disorders were in fluenced by shared genetic factors (rg= 0.38– 0.56), with all genetic overlap between substances attributable to genetic in fluences shared with conduct disorder. Genes in fluencing individual sub-stance use disorders were also signi ficant, explaining 40– 73% of the genetic variance per substance
There is substantial evidence that similar genetic fac-tors in fl uence liability to multiple substance use disor-ders (e.g. Kendleret al. 2003a , 2007; Rhee et al. 2006; Young et al. 2006; Xian et al. 2008; Sartor et al. 2010; Palmeret al.2013)
results indicating the absence of a residual genetic correlation between alcohol and cannabis dependence when accounting for genetic overlap with antisocial personality disorder have also been noted in adult Vietnam era males (Fu et al. 2002). Other studies suggest that this shared liability extends to additional aspects of externalizing behavior, including novelty seeking and non-substance-related behavioral disinhibition (Krueger et al. 2002; Hicks et al.2011)
Alcohol initiation was endorsed by almost 99% of the respondents, with 89% of the sample reporting having consumed at least fi ve drinks in a 24-h period and 26% of those having consumed 55 drinks in a 24-h period reporting a lifetime history of AD. Over 88% of respondents had tried cigarettes, with 33% of initiators having a history of ND. Cannabis initiation was endorsed by 63% of respon-dents, with 28% of initiators having a history of CAD
the genetic correlation across substances was entirely attributable to genes shared with conduct disorder
Kendler et al .( 2007) noted that substance-specifi c genetic variance ranged from 3% for cocaine to 91% for caffeine abuse/dependence symptoms
Our results are consistent with previous reports (Fu et al. 2002; Kendler et al. 2003b ; Button et al. 2006; Hicks et al. 2011) showing that alcohol and drug de-pendence tend to share an appreciable proportion of their genetic liability with conduct and antisocial per-sonality disorder
a signifi cant proportion of the genetic variance in ND (40%), AD (73%), and CAD (47%) was substance-speci fi c
Greer G, Grob C, Halberstadt A. 2014 “PTSD symptom reports of patients evaluated for the New Mexico medical cannabis program” Journal of Psychoactive Drugs 46(1): 73-77
http://www.tandfonline.com/doi/full/10.1080/02791072.2013.873843
Greater than 75% reduction in CAPS symptom scores were reported when patients were using cannabis compared to when they were not
In one unpublished, open-label pilot study, smoked medical cannabis containing 23% tetrahydrocannabinol (THC) and less than 1% cannabidiol was administered to 29 male Israeli combat veterans with PTSD, with instructions to smoke it daily (Mashiah 2012). The baseline score on the Clinician Administered Posttraumatic Scale for DSM-IV (CAPS) was 98 for the entire group, and post-treatment scores in three subgroups after four to 11 months of treatment ranged from 54 to 60
Criterion B (core symptom cluster of re-experiencing), which decreased from 29.5 ± 6.4 to 7.3 ± 5.9 (F(1,79) = 734.98, p < 0.0001); Criterion C (numbing and avoidance), which decreased from 38.2 ± 8.4 to 8.7 ± 8.0 (F(1,79) = 783.73, p < 0.0001); and Criterion D (hyperarousal), which decreased from 31.0 ± 6.2 to 6.6 ± 6.0 (F(1,79) = 910.79,p < 0.0001)
Some degree of intentional or unintentional exaggeration of symptom differences on the part of the patients is likely, and some unintentional bias on the part of the psychiatrist conducting the evaluations is possible
variability in scores with cannabis use was relatively high, with the standard deviation being almost equal to the mean total scores and the scores of the three symptom clusters. If patients had consistently reported frequent and severe symptoms without cannabis and almost no symptoms with cannabis in order to make sure they qualified for the Program, one would expect less variability in the cannabis scores. Finally, the relatively consistent reporting of low or “0” scores on Criterion C3 without cannabis (see ) is another indication that most patients were not malingering by exaggerating their no-cannabis scores for every single symptom in order to qualify for the program
Because only patients who reported benefit from cannabis in reducing their PTSD were studied, no conclusions can be drawn as to what proportion or type ofPTSD patients would benefit from treatment with cannabis or its constituents
Grella C, Joshi V. 2003. “Treatment processes and outcomes among adolescents with a history of abuse who are in drug treatment” Child Maltreat 8(1): 7-18
http://www.ncbi.nlm.nih.gov/pubmed/12568501
N = 803, physical and/or sexual abuse was reported by 59% of the girls and 39% of the boys; physically abused adolescents less likely to stay abstinent after treatment
Griffith-Lendering M, Wigman J, Prince van Leeuwen A, Huijbregts S, Huizink A, Ormel J, Verhulst F, van Os J, Swaab H, Vollebergh W. 2013. “Cannabis use and vulnerability for psychosis in earl adolescence–a TRAILS study” Addiction 108(4): 733-40
http://www.ncbi.nlm.nih.gov/pubmed/23216690 add to dropbox
A large prospective population study of Dutch adolescents [the TRacking Adolescents’ Individual Lives Survey (TRAILS) study].Participants A total of 2120 adolescents with assessments at (mean) age 13.6, age 16.3 and age 19.1
ignificant associations (r = 0.12–0.23) were observed between psychosis vulnerability and cannabis use at all assessments. Also, cannabis use at age 16 predicted psychosis vulnerability at age 19 (Z = 2.6, P < 0.05). Further-more, psychosis vulnerability at ages 13 (Z = 2.0, P < 0.05) and 16 (Z = 3.0, P < 0.05) predicted cannabis use at, respectively, ages 16 and 19
there is increasing evidence for stability or continuity (from adoles-cence onwards) of psychosis symptoms [23–25]
confounding variables such as socio-economic status (SES), parental psychopathology and use of other substances
exclusion criteria were: (i) adolescent’s inability of participating because of intellectual disability or a serious physical illness or handicap; and (ii) Dutch-speaking parent or parent surrogate not available, and not feasible to administer a part of the measurements in the parent’s own language. Of all subjects who were approached (n = 3145), 6.7% were excluded
Using categories no use (i), one to two times (ii), three to six times (iii) and seven times or more (v) resulted in the best Tukey solutions for T2, T3 and T4
Vulnerability for psychosis at T2 predicted cannabis use at T3… cannabis use measured at T2 did not predict psychosis vulnerability measured at T3
vulnerability for psychosis at T3 predicted cannabis use at T4… cannabis use measured at T3 significantly predicted psy-chosis vulnerability measured at T4
With respect to self-medication, it has been hypothesized specifically that individuals with psychosis symptoms use cannabis to improve their mood or to control one’s feelings, boredom, social motives, improving sleep, anxiety and agitation, although some studies indicate that individuals with psy-chosis symptoms use cannabis for similar reasons as the general population as well, i.e. ‘to get high’, relax and have fun [10,11,46]
a cascading model would fit the temporal associations between cannabis use and psy-chosis vulnerability best, particularly because associations between cannabis use and psychosis vulnerability became stronger at later assessments
it may be argued that the choice of potential confounders could have been more refined. For example, the genetic variation associated with psychosis vulnerability and substance use could manifest itself in (impairments in) certain (dopamine or serotonin-dependent) cognitive abilities that could have been assessed as well
Gruber A, Pope H. 2002. “Marijuana use among adolescents” Pediatr Clin North Am 49(2): 389-413
http://www.ncbi.nlm.nih.gov/pubmed/11993290
More than half of US adolescents will experiment with marijuana. Of those who try marijuana more than once, approximately one third will subsequently use marijuana regularly, although most will have stopped by their late 20s.
Gruber S, Yurgelun-Todd D. 2005. “Neuroimaging of marijuana smokers during inhibitory processing: a pilot investigation” Brain Res Cogn Brain Res 23(1):107-18
http://www.ncbi.nlm.nih.gov/pubmed/15795138
chronic marijuana abusers in Stroop demonstrated significantly lower anterior cingulate activity in focal areas of the anterior cingulate cortex and higher midcingulate activity relative to controls, although both groups were able to perform the task within normal limits. Normal controls also demonstrated increased activity within the right dorsolateral prefrontal cortex (DLPFC) during the interference condition, while marijuana smokers demonstrated a more diffuse, bilateral pattern of DLPFC activation
DTI measures in frontal regions, which include the genu and splenium of the corpus callosum and bilateral anterior cingulate white matter regions, showed no between group differences in fractional anisotropy (FA), a measure of directional coherence within white matter fiber tracts, but a notable increase in trace, a measure of overall isotropic diffusivity in marijuana smokers compared to controls
Gruber S, Rogowska J, Yurgelun-Todd D. 2009. “Altered affective response in marijuana smokers: an FMRI study” Drug Alcohol Depend 105: 139-153
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2752701/
Despite no between-group differences on clinical or demographic measures, smokers demonstrated a relative decrease in both anterior cingulate and amygdalar activity during masked affective stimuli compared to controls, who showed relative increases in activation within these regions during the viewing of masked faces
none of the chronic, heavy marijuana subjects were acutely intoxicated or “high” at the time of the scan and had been abstinent for a minimum of 12–16 hours, and are therefore not likely to reflect acute effects of the drug
Gruber S, Silveri M, Dahlgren M, Yurgelun-Todd. 2011. “Why so impulsive? White matter alterations are associated with impulsivity in chronic marijuana users” Exp Clin Psychopharmacol 19: 231-242
http://www.ncbi.nlm.nih.gov/pmc/articles/pmid/21480730/
significant reductions in left frontal FA in MJ smokers associated with impulsivity
Fifteen adult chronic, heavy MJ smokers who had smoked at least 3,000 joints in their lifetime, average age of onset of 14.9 years, smoked an average of 25.5 joints per week, used at least four of last 7 days and tested positive for cannabinoids and 15 non-MJ smoking control subjects who were age, sex, and education matched; twice as much alcohol use in MJ group (9.6 days in last month compared to 4)
Axis I pathology excluded by Structured Clinical Interview for DSM-IV
Gruber S, Sagar K, Gahlgren M, Racine M, Lukas S. 2012. “Age of onset of marijuana use and executive function” Psychology of Addictive Behaviors 26(3): 496-506
http://psycnet.apa.org/?&fa=main.doiLanding&doi=10.1037/a0026269
Thirty-four chronic, heavy MJ smokers separated into early and late onset groups, and 28 non-MJ smoking controls completed a battery of neurocognitive measures
As hypothesized, MJ smokers performed more poorly than controls on several measures of executive function. Age of onset analyses revealed that these between-group differences were largely attributed to the early onset group, who were also shown to smoke twice as often and nearly 3 times as much MJ per week relative to the late onset smokers
Exposure to MJ during a period of neurodevelopmental vulnerability, such as adolescence, may result in altered brain development and enduring neuropsychological changes
Gruber S, Dahlgren M, Sagar K, Gonenc A, Lukas S. 2014. “Worth the wait: effects of age of onset of marijuana use on white matter and impulsivity” Psychopharmacology (Berl) 321(8): 1455-1465
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3967072/
Twenty-five MJ smokers and 18 healthy controls… MJ smokers had to have reported smoking a minimum of 2,500 times, used MJ at least 5 out of the last 7 days, tested positive for urinary cannabinoids, and met DSM-IV criteria for MJ abuse or dependence
divided into early onset (regular use prior to age 16) and late onset (age 16 or later) groups
MJ smokers exhibited significantly reduced fractional anisotropy (FA) relative to controls, as well as higher levels of impulsivity. Earlier MJ onset was also associated with lower levels of FA. Interestingly, within the early onset group, higher impulsivity scores were correlated with lower FA, a relationship that was not observed in the late onset smokers
no Axis I pathology was present other than MJ use in the smoking group
Comparisons of early and late onset smokers revealed no significant differences in patterns of MJ use between the two groups. However, it is of note that early onset smokers smoked slightly more often (smokes/week; 18.76 vs. 15.51) and more than twice as much MJ per week (grams/week; 14.65 vs. 6.66) as late onset smokers [MV: how the fuck is 14.65 grams per week not significantly different than 6.66 grams per week?]
when the MJ-smoking group was separated into those with early vs. late onset, it appeared that these relationships were driven almost exclusively by the early onset smokers, as higher impulsivity scores on all BIS subscales were significantly associated with lower FA in both the left and right genu of early onset smokers
decreased white matter integrity is associated with higher levels of impulsivity, specifically among individuals who initiated MJ use prior to age 16
higher concentrations of cannabinoid receptors in neural fiber tracts during adolescence may therefore represent a period of specific vulnerability to the effects of MJ use on white matter microstructure
Study findings may therefore be specific to individuals who do not endorse the more negative effects of marijuana use (i.e., psychological issues, inability to stop or cut down on use, withdrawal effects) and to those who do not meet for dependence, despite frequent, heavy use
it remains unclear whether the reductions in FA in MJ smokers noted in the present study are the result of demyelination or damage to white matter, delayed or altered brain developmental patterns in MJ smokers, and if they precede or are the result of MJ use
Gunduz-Cinar O, MacPherson K, Cinar R, Gamble-George J, Sugden K, Williams B, Godlewski G, Ramikie T, Gorka A, Alapafuja S, Nikas S, Makriyannis A, Poulton R, Patel S, Hariri A, Caspi A, Moffitt T, Kunos G, Holmes A. 2013. “Convergent translational evidence of a role for anandamide in amygdala-mediated fear extinction, threat processing and stress-reactivity.” Molecular Psychiatry 18(7): 813-23.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3549323/
FAAH inhibition reduces fear in rats given extinction training; effect prevented by injection of CB1 antagonist. Found genetic variances in FAAH influence stress-reactivity
Gupta M. 2013. “Review of somatic symptoms in post-traumatic stress disorder” Int Rev Psychiatry 25(1): 86-99
http://www.ncbi.nlm.nih.gov/pubmed/23383670
Post-traumatic stress disorder (PTSD) is associated with both (1) ‘ill-defined’ or ‘medically unexplained’ somatic syndromes, e.g. unexplained dizziness, tinnitus and blurry vision, and syndromes that can be classified as somatoform disorders (DSM-IV-TR); and (2) a range of medical conditions, with a preponderance of cardiovascular, respiratory, musculoskeletal, neurological, and gastrointestinal disorders, diabetes, chronic pain, sleep disorders and other immune-mediated disorders
cardiovascular disease, especially hypertension, and immune-mediated disorders
PTSD is associated with limbic instability and alterations in both the hypothalamic- pituitary-adrenal and sympatho-adrenal medullary axes, which affect neuroendocrine and immune functions, have central nervous system effects resulting in pseudo-neurological symptoms and disorders of sleep-wake regulation, and result in autonomic nervous system dysregulation. Hypervigilance, a central feature of PTSD, can lead to ‘local sleep’ or regional arousal states, when the patient is partially asleep and partially awake, and manifests as complex motor and/or verbal behaviours in a partially conscious state
Gur R. 1999. “Is schizophrenia a lateralized brain disorder?” Schizophrenia Bulletin 25: 7-10
http://schizophreniabulletin.oxfordjournals.org/content/25/1/7.abstract
laterality hypotheses of schizophrenia were proposed more than two decades ago (Gruzelier and Venables 1974; Flor-Henry 1976; Gur 1977, 1978) and have since been tested using increasingly powerful neurobehavioral, neuroanatomic, and neurophysiologic methods
the question of whether schizophrenia is characterized by dysfunction of one or the other hemisphere is simplistic and needs to be reformulated
Patients with schizophrenia from a large prospective sample show meager indication of lateralized disturbances in sensory processing and motor speed. They do, however, have more language than spatial deficits, implicating relative left hemispheric dysfunction in regions needed for complex cognitive behavior. Laterality effects in performance were mediated by sex differences, and the results underscore the importance of examining sex differences in laterality research
One may vacillate, after reading this diverse set of contributions, between being overwhelmed by the amount of data that have converged on the issue of laterality in schizophrenia and feeling some exasperation at the paucity of solid answers for questions that seem quite rudimentary
Haj-Dahmane S, Shen R. 2014. “Chronic stress impairs α1-adrenoceptor-induced endocannabinoid-dependent synaptic plasticity in the dorsal raphe nucleus” Journal of Neuroscience 34(44): 14560-14570
http://www.jneurosci.org/content/34/44/14560.short
Alpha 1-adrenergic receptors (α1-ARs) control the activity of dorsal raphe nucleus (DRn) serotonin (5-HT) neurons and play crucial role in the regulation of arousal and stress homoeostasis
in the control condition, activation of α1-ARs induced an inward current and long-term depression (LTD) of glutamate synapses of DRn 5-HT neurons. The α1-AR LTD was initiated by postsynaptic α1-ARs but mediated by a decrease in glutamate release. The presynaptic expression of the α1-AR LTD was signaled by retrograde endocannabinoids (eCBs)
chronic exposure to restraint stress profoundly reduced the magnitude of α1-AR LTD but had no effect on the amplitude of α1-AR-induced inward current. Chronic restraint stress also reduced the CB1 receptor-mediated inhibition of EPSC and the eCB-mediated depolarization-induced suppression of excitation. Collectively, these results indicate that chronic restraint stress impairs the α1-AR LTD by reducing the function of presynaptic CB1 receptors and reveal a novel mechanism by which noradrenaline controls synaptic strength and plasticity in the DRn. They also provide evidence that chronic stress impairs eCB signaling in the DRn, which may contribute, at least in part, to the dysregulation of the stress homeostasis
Hamilton I, Lloyd C, Hewitt C, Godfrey C. 2013. “Effect of reclassification of cannabis on hospital admissions for cannabis psychosis: a time series analysis” International Journal of Drug Policy 25(1): 151-156
http://www.ijdp.org/article/S0955-3959(13)00090-X/abstract
There was a significantly increasing trend in cannabis psychosis admissions from 1999 to 2004. However, following the reclassification of cannabis from B to C in 2004, there was a significant change in the trend such that cannabis psychosis admissions declined to 2009. Following the second reclassification of cannabis back to class B in 2009, there was a significant change to increasing admissions.
Hampson AJ, Grimaldi M, …, Wink D. 1998. “Cannabidiol and delta-9-tetrahydrocannabinol are neuroprotective antioxidants.” Proc Natl Acad Sci. 95(14):8268-8273
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC20965/
Han J, Liu Z, Ren W, Zhang X. 2011. “Counteractive effects of cannabinoid and nicotine-addictive behavior” Neuroreport 22(4): 181-4
http://www.ncbi.nlm.nih.gov/pubmed/21304327
Our recent results suggest that cannabinoid exposure induces conditioned place preference (CPP) through facilitated induction of synaptic long-term depression at dopamine circuitry of the midbrain ventral tegmental area (VTA). Here, we show that chronic nicotine exposure also induces CPP, but facilitates the induction of synaptic long-term potentiation in the VTA. Coadministration of cannabinoid and nicotine leads to a blockade of facilitated long-term depression and long-term potentiation induction in these neurons and elimination of CPP. These findings point to counteractive effects of cannabinoid and nicotine-addictive behavior through opposite changes in synaptic plasticity of dopamine circuitry of the VTA
Haughey H, Marshall E, Schacht J, Louis A, Hutchison K. 2008. “Marijuana withdrawal and craving: influence of the cannabinoid receptor 1 (CNR1) and fatty acid amide hydrolase (FAAH) genes” Addiction 103(10): 1678-1686
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2873690/
Glutamate toxicity reduced by cannabidiol and THC; also were shown to prevent hydroperoxide-induced oxidative damage as well as or better than other antioxidants in a chemical (Fenton reaction) system and neuronal cultures. Cannabidiol was more protective against glutamate neurotoxicity than either ascorbate or α-tocopherol, indicating it to be a potent antioxidant. may be useful for treatment of oxidative neurological disorders such as cerebral ischemia. also protective against calcium-signalling mediated forms of neurotoxicity
Heifets B, Castillo P. 2009. “Endocannabinoid signaling and long-term synaptic plasticity” Annual Review of Physiology 71: 283-306
http://www.ncbi.nlm.nih.gov/pubmed/19575681 – in dropbox
we review the major molecular and cellular mechanisms underlying eCB-LTD, as well as the potential physiological relevance of this widespread form of synaptic plasticity
Activation of the CB1R, and subsequent long-term reduction of transmitter release at the same synapse, defines eCB-LTD
CB1Rs can engage a wide range of effector molecules, in-cluding (but not limited to) voltage-dependent Ca2+ channels (VDCCs), K+ channels, pro-tein kinase A (PKA), and mitogen-activated protein kinase (MAPK) (recently reviewed in Reference 14)
eCB-LTD induction typically begins with a transient increase in activity at glutamatergic afferents and a concomitant release of eCBs from a target (postsynaptic) neuron (Figure 1 ). eCBs then travel backward (retrogradely) across the synapse, activating CB1Rs on the presy-naptic terminals of either the original afferent (homosynaptic eCB-LTD) or nearby afferents (heterosynaptic eCB-LTD
In addition to mGluR-I, metabotropic dopamine (type 2; D2), muscarinic acetylcholine (type 1/3; M 1 /M3), serotonin (type 2; 5-HT2), orexin, and chole-cystokinin receptors are all effective stimuli for eCB release… D2 stimulate the formation of AEA rather than 2-AG… are required for corticostriatal eCB-LTD… a role for AEA in this process
D2Rs may directly stimulate eCB release from medium spiny neurons, or, as recently suggested, D2Rs may exert an indirect effect on eCB release via cholinergic interneurons in dorsal striatum. However, a recent study has shown that the presence or absence of D2Rs on MSNs determines whether eCB-LTD can be induced at an afferent glutamatergic synapse, owing to a D2R-mediated facilitation of eCB release
the requirement for coincident activation of CB1Rs and presynaptic activity provides an additional mechanism to ensure synapse specificity such that only active fibers undergo eCB-LTD
Sj ¨ostrom et al. (26) postulated that coincident activa-tion of presynaptic NMDA and CB1 receptors is required for the induction of eCB-t-LTD.
CB1R agonists alone do not trigger long-term plasticity without co-incident presynaptic activity
heterosynap-tic eCB-LTD at excitatory synapses in the im-mature hippocampus is likely due to a CB1R-dependent reduction of presynaptic excitability through activation of K+ channels at presynaptic fibers
Interestingly, brain-derived neurotrophic factor (BDNF), which may be re-leased from strongly activated presynaptic sites, appears to prevent the induction of homosy-naptic eCB-LTD. presumably, an interaction between the CB1R and TrkB (the high-affinity receptor for BDNF) at active presynaptic terminals leads to the blockade of CB1R signal-ing
LTD is a leading model to explain the synap-tic weakening that occurs at cortical synapses after a period of sensory deprivation
Long-term changes in synaptic strength are also believed to underlie associative memory forma-tion in hippocampus and amygdala… eCBs and CB1Rs were specifically required for extinguishing an established fear or spatial memory but were not needed to acquire these types of memories
Heinrichs R, Zakzanis K. 1998. “Neurocognitive deficit in schizophrenia: a quantitative review of the evidence” Neuropsychology 12(3): 426-45
http://www.ncbi.nlm.nih.gov/pubmed/9673998/
Moderate to large raw effect sizes (d > .60) were obtained for all 22 neurocognitive test variables, and none of the associated confidence intervals included zero. The results indicate that schizophrenia is characterized by a broadly based cognitive impairment, with varying degrees of deficit in all ability domains measured by standard clinical tests
de Heredia F, Gomez-Martinez S, Marcos A. 2012. “Obesity, inflammation and the immune system” Proc Nutr Soc 71(2): 332-8
http://www.ncbi.nlm.nih.gov/pubmed/22429824
Obesity shares with most chronic diseases the presence of an inflammatory component, which accounts for the development of metabolic disease and other associated health alterations… Obesity, like other states of malnutrition, is known to impair the immune function, altering leucocyte counts as well as cell-mediated immune responses… an altered immune function contributes to the pathogenesis of obesity… there is a positive feedback loop between local inflammation in adipose tissue and altered immune response in obesity, both contributing to the development of related metabolic complications
Hindocha C, Shaban N, Freeman T, Das R, Gale G, Schafer G, Falconer C, Morgan C, Curran H. 2015. “Associations between cigarette smoking and cannabis dependence: a longitudinal study of young cannabis uers in the United Kingdom” Drug Alcohol Depend 148: 165-171
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4337852/
Participants were 298 cannabis and tobacco users aged between 16 and 23; follow-up consisted of 65 cannabis and tobacco users
Regression analyses at baseline showed cigarette smoking (frequency of cigarette smoking: B = 0.029, 95% CI = 0.01, 0.05; years of cigarette smoking: B = 0.159, 95% CI = 0.05, 0.27) accounted for 29% of the variance in cannabis dependence when controlling for frequency of cannabis use. At follow-up, only baseline cannabis dependence predicted follow-up cannabis dependence (B = 0.274, 95% CI = 0.05, 0.53). At baseline, cigarette smoking mediated the relationship between frequency of cannabis use and dependence (B = 0.0168, 95% CI = 0.008, 0.288) even when controlling for possible confounding variables (B = 0.0153, 95% CI = 0.007, 0.027)
As nicotine is more addictive than cannabis, tobacco smoking may be a primary driver of continued use and relapse in co-dependent users.
About 90% of cannabis users also identify as cigarette smokers (Agrawal et al., 2012), however, this exists as a complicated relationship given that increased cigarette smoking may substitute for reduced cannabis consumption (Allsop et al., 2014) and vice versa. Users of both drugs report more severe symptoms of CUD (Peters et al., 2012). Half of adults seeking treatment for CUD also smoke cigarettes and treatment outcomes for those using both cannabis and tobacco, in comparison to cannabis alone, are poor (Agrawal et al., 2009). Moreover, relative to those with a CUD, those with co-occurring nicotine dependence show poorer psychiatric and psychosocial outcomes (Peters et al., 2014; Ramo et al., 2013). In a recent controlled laboratory study, Haney et al. (2012) found that the strongest predictor of relapse in cannabis dependent individuals was their cigarette smoking status
Cigarette smoking at baseline was predictive of CUD at baseline when controlling for cannabis use variables in young people who smoke cannabis and tobacco. The most efficient model accounted for 30% of the variance in cannabis dependence which is considered to be a large effect size as R > 0.5 (Cohen, 1988). However, this seems no longer the case four years later, where only baseline CUD predicted follow-up CUD… cigarette smoking may enhance the dependence-forming effects of cannabis. Alternatively, our results may suggest that CUD (as measured by the SDS) may capture some aspects of nicotine dependence in a subset of young people with CUD
days of cannabis use per month at baseline were not associated with cannabis dependence at follow up
Ho B-C, Andreasen N, Ziebell S, Pierson R, Magnotta V. 2011. “Long-term antipsychotic treatment and brain volumes” Arch Gen Psychiatry 68(2): 128-137
http://archpsyc.jamanetwork.com/article.aspx?articleid=211084
During longitudinal follow-up, antipsychotic treatment reflected national prescribing practices in 1991 through 2009. Longer follow-up correlated with smaller brain tissue volumes and larger cerebrospinal fluid volumes… More antipsychotic treatment was associated with smaller gray matter volumes. Progressive decrement in white matter volume was most evident among patients who received more antipsychotic treatment. Illness severity had relatively modest correlations with tissue volume reduction, and alcohol/illicit drug misuse had no significant associations when effects of the other variables were adjusted.
Hoffman A, Oz M, Yang R, Lichtman A, Lupica C. 2007. “Opposing actions of chronic Δ9-tetrahydrocannabinol and cannabinoid antagonists on hippocampal long-term potentiation” Learn Mem 14(1-2): 63-74
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1828281/
Although cannabinoids acutely reduce glutamate release and block hippocampal long-term potentiation (LTP), a potential substrate for learning and memory, the consequences of prolonged exposure to Δ9-THC for hippocampal function are poorly understood
rats
Δ9-THC also blocked HFS-LTP after 3 d, but not 1 d of treatment. The complete blockade of LTP persisted for 3 d after the last Δ9-THC injection, and full reversal of the LTP deficit was not observed up to 14 d following Δ9-THC withdrawal. The cannabinoid antagonist AM251 (2 mg/kg), administered before each Δ9-THC injection prevented the blockade of LTP, and 7-d treatment with AM251 alone significantly increased the level of LTP. Chronic Δ9-THC also produced tolerance to the inhibition of synaptic GABA, but not glutamate release by the agonist WIN55,212-2
cannabinoid-induced impairment of hippocampal LTP was reversed via the facilitation of NMDA receptor function by either the depolarization of CA1 pyramidal neurons or by removing Mg2+ from the extracellular space (Misner and Sullivan 1999). This suggested that the blockade of LTP by cannabinoid agonists resulted from a decrease in the probability of glutamate release via presynaptic receptors (Misner and Sullivan 1999)
the relationship between presynaptic axon fiber volley and fEPSP amplitudes in slices obtained from 7-d vehicle- and 7-d Δ9-THC-treated rats was examined. There was no effect of chronic Δ9-THC on this relationship (), suggesting that synaptic strength was not altered by the chronic treatment.
the consequences of chronic Δ9-THC exposure were not mediated by ongoing pharmacological actions of this cannabinoid agonist following its accumulation in tissue, but rather through more enduring neurobiological changes in hippocampal function brought about through repeated exposure to the drug
cannabinoid receptor antagonists do not acutely alter HFS-LTP, and that endogenous cannabinoids released during HFS (Stella et al. 1997) are not likely involved in the initiation or maintenance of HFS-LTP
Chronic exposure to Δ9-THC does not alter NMDA receptor function, but reduces paired pulse facilitation of AMPA-mediated EPSCs
in contrast to GABAergic synapses (see below), the inhibitory effect of WIN55,212-2 on glutamatergic fEPSPs was not altered by chronic Δ9-THC
Harder V, Morral A, Arkes J. 2006. “Marijuana use and depression among adults: Testing for causal associations” Addiction 101(10): 1463-72
http://www.ncbi.nlm.nih.gov/pubmed/16968348
marijuana users 1.4 times as likely to be depressed. Dropped to 1.1 after adjustment for risk factors
Harder V, Stuart E, Anthony J. 2008. “Adolescent cannabis problems and young adult depression: male-female stratified propensity score analyses” Am J Epidemiol 168(6): 592-601
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2727198/
Cannabis use and depression are two of the most prevalent conditions worldwide
mid-Atlantic metropolitan area of the United States, repeated assessments from 1985 (at age 6 years) through 2002 (at age 21 years) were made for 1,494 individuals (55% female)
The estimated risk of young adult depression for adolescents with cannabis problems was not significantly different from that for comparison adolescents for either females (odds ratio = 0.7, 95% confidence interval: 0.2, 2.3) or males (odds ratio = 1.7, 95% confidence interval: 0.8, 3.6). The evidence does not support a causal association linking adolescent-onset cannabis problems with young adult depression
Cannabis problem users (the “exposed” group) are different from comparison individuals on many measured preexposure covariates… a higher percentage of the cannabis problem users were daily tobacco users, had problem alcohol use, or had slightly higher concentration and behavior problems than the comparison individuals
Our study does not support the hypothesis that adolescent-onset cannabis problem use causes young adult depression. Two other causal hypotheses remain: 1) Depression causes individuals to manage their symptoms through self-medication by use of cannabis, and 2) a common genetic or environmental influence causes both depression and cannabis use. Although there is consistent evidence that depression does not cause cannabis use (50), there is evidence in support of the common cause hypothesis through the use of co-twin methodology to control for genetic influences (51)
Hariri A, Gorka A, Hyde L, Kimak M, Halder I, Ducci F, Ferrell R, Goldman D, Manuck S. 2009. “Divergent Effects of Genetic Variation in Endocannabinoid Signaling on Human Threat- and Reward-Related Brain Function” Biol Psychiatry 66(1): 9-16
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3215587/?report=classic
Carriers of FAAH 385A, associated with reduced enzyme and, possibly, increased eCB signaling, had decreased threat-related amygdala reactivity but increased reward-related ventral striatal reactivity in comparison to C385 homozygotes. Similar divergent effects of FAAH C385A genotype were manifest at the level of brain-behavior relationships. 385A carriers showed decreased correlation between amygdala reactivity and trait anxiety but increased correlation between ventral striatal reactivity and delay discounting, an index of impulsivity.
Harkness K, Bruce A, Lumley M. 2006. “The role of childhood abuse and neglect in the sensitization to stressful life events in adolescent depression” J Abnorm Psychol 115(4): 730-41
http://www.ncbi.nlm.nih.gov/pubmed/17100530/
Consistent with hypotheses, adolescents with a history of childhood abuse and/or neglect reported a lower level of threat of stressful life events prior to episode onset than that reported by those without… his effect was robust when controlling for level of chronic difficulties, which was higher in those with childhood abuse and/or neglect. The authors suggest that childhood abuse and/or neglect may be an important risk factor that sensitizes individuals to the effects of acute independent life events
Harrison G, Gunnell D, Glazebrook C, Pake K, Kwiecinski R. 2001. “Association between schizophrenia and social inequality at birth: case-control study” British Journal of Psychiatry 179: 346-350
http://bjp.rcpsych.org/content/179/4/346.full
Those both born in “deprived area” and to social class IV-V father had odds ratio of 8.1 for developing schizophrenia
Harrow M, Jobe T. 2013. “Does long-term treatment of schizophrenia with antipsychotic medications facilitate recovery?” Schizophr Bull 39(5): 962-965
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3756791/
Antipsychotic medications are viewed as cornerstones for both the short-term and long-term treatment of schizophrenia
antipsychotic medications are viewed as the cornerstone of treatment, in both the short-term and the long-term treatments of patients with schizophrenia
Prolonged use of antipsychotic medications is viewed as a key factor in treatment for schizophrenia, but there is very little systematic evidence for the long-term benefits of antipsychotics. There is even some longitudinal data suggesting the opposite
within the first 6–10 months after discontinuation, 25%–55% of schizophrenia patients discontinued from antipsychotics relapse
patients with schizophrenia not on antipsychotics for a prolonged period do not show this tendency to relapse when they remain unmedicated
when vulnerable patients are treated with antipsychotics for a prolonged period, this increases their chances of relapsing if they subsequently discontinue
The discontinuation effect includes the potential of medication-generated buildup, prior to discontinuation, of supersensitive dopamine receptors, or the buildup of excess dopamine receptors, or supersensitive psychosis, as indicated by multiple studies by Seeman14 and others13 of dopamine-blocking agents using animal models
“breakthrough supersensitivity during ongoing antipsychotic treatment undermines treatment efficacy.”
“… nothing is known about the effects of antipsychotic drugs compared to placebo after three years.”
a sample of schizophrenia patients who were treated continuously with antipsychotics over 15-year and 20-year periods have shown considerable psychopathology and few sustained periods of recovery
the sample of schizophrenia patients who were untreated for many years showed significantly better outcomes than did those on antipsychotics
Other longitudinal studies have found similar results. This includes important longitudinal studies such as the Vermont studies of C. Harding20,21 and the Chestnut Lodge Study.22 In Canada, the Alberta Hospital Studies of Bland (1978) found similar results. Overseas the longitudinal studies of M. Bleuler (1978) led to his commenting about relapses for many schizophrenia patients treated with antipsychotics. The important World Health Organization (WHO) Study and the Determinants of Outcome of Severe Mental Disorders (DOSMED) Study by Edgerton and Cohen found better outcomes in many developing countries where only a small percentage of schizophrenia patients were treated with antipsychotics
Other evidence, also from longitudinal studies, suggest that long-term outcome for schizophrenia in the modern era is not much better than it was 60–80 years ago.23,24 In regard to relapses, our own evidence indicates that many schizophrenia patients who have not been treated with antipsychotic medications for prolonged periods show a low rate of relapse over the next 5-year period
Harley M, Kelleher I, Clarke M, Lynch F, Arseneault L, Connor D, Fitzpatrick C, Cannon M. 2010. “Cannabis use and childhood trauma interact additively to increase the risk of psychotic symptoms in adolescence” Psychological Medicine 40: 1627-1634
http://www.ncbi.nlm.nih.gov/pubmed/19995476 in dropbox
both cannabis use and childhood trauma were significantly associated with risk of experiencing psychotic symptoms. However, the presence of both childhood trauma and early cannabis use significantly increased the risk for psychotic symptoms beyond the risk posed by either risk factor alone, indicating that there was a greater than additive interaction between childhood trauma and cannabis use
childhood trauma was reported in 11.3% of participants interviewed. Those adolescents who had experienced childhood trauma were almost five times more likely to use cannabis… also five times more likely to develop psychotic symptoms
Hart C, van Gorp W, Haney M, Foltin R, Fischman M. 2001. “Effects of acute smoked marijuana on complex cognitive performance” Neuropsychopharmacology 25: 757-765
http://www.nature.com/npp/journal/v25/n5/full/1395716a.html
Although the ability to perform complex cognitive operations is assumed to be impaired following acute marijuana smoking, complex cognitive performance after acute marijuana use has not been adequately assessed under experimental conditions… Although marijuana significantly increased the number of premature responses and the time participants required to complete several tasks, it had no effect on accuracy on measures of cognitive flexibility, mental calculation, and reasoning… acute marijuana smoking produced minimal effects on complex cognitive task performance in experienced marijuana users
Marijuana increased premature responding and the amount of time needed to complete several MicroCog™ tasks, including the immediate recall, tic-tac, clocks, and mental calculation tasks. Nevertheless, accuracy on these and other cognitive tasks were unaltered after marijuana administration
psychomotor performance was only subtly affected by smoked marijuana: following high Δ9-THC concentration cigarettes, participants exhibited a decrement in performance only during the immediate recall task, while performance on a tracking task was markedly improved
The most salient cognitive effect from the present investigation was the finding that premature responses were increased after volunteers smoked the high Δ9-THC concentration cigarette
more complex cognitive performance is only minimally affected following acute marijuana smoking.
heavy marijuana smokers show less cognitive impairment following acute marijuana administration (Meyer et al. 1971; Rickles et al. 1973;Cohen and Rickles 1974)
marijuana use history plays less of a role in subjective ratings and heart rate responses than it does on cognitive performance after smoking a single marijuana cigarette
Hart C, Ilan A, Gevins A, Gunderson E, Role K, Colley J, Foltin R. 2010. “Neurophysiological and cognitive effects of smoked marijuana in frequent users” Pharmacol Biochem Behav 96(3): 333-41
http://www.ncbi.nlm.nih.gov/pubmed/20600251
electroencephalographic (EEG) signals were recorded as daily marijuana users performed additional tests of immediate working memory and delayed episodic memory, before and after smoking marijuana
Overall performance accuracy was not significantly altered by marijuana, although the drug increased response times during task performance and induced a response bias towards labeling “new” words as having been previously seen in the verbal episodic memory task. Marijuana reduced slow wave evoked potential amplitude in the episodic memory task and decreased P300 amplitude and EEG power in the alpha band in the spatial working memory task
van der Hart O, Nijenhuis E, Steele K. 2005. “Dissociation: an insufficiently recognized major feature of complex posttraumatic stress disorder” J Trauma Stress 18(5): 413-23
http://www.ncbi.nlm.nih.gov/pubmed/16281239/
The role of dissociation in (complex) posttraumatic stress disorder (PTSD) has been insufficiently recognized for at least two reasons: the view that dissociation is a peripheral, not a central feature of PTSD, and existing confusion regarding the nature of dissociation… traumatization essentially involves some degree of division or dissociation of psychobiological systems that constitute personality. One or more dissociative parts of the personality avoid traumatic memories and perform functions in daily life, while one or more other parts remain fixated in traumatic experiences and defensive actions. Dissociative parts manifest in negative and positive dissociative symptoms that should be distinguished from alterations of consciousness. Complex PTSD involves a more complex structural dissociation than simple PTSD
Hartung B, Schwender H, Roth E, Hellen F, Mindiashvili N, Rickert A, Ritz-Timme S, Grieser A, Monticelli F, Daldrup T. 2015. “The effect of cannabis on regular cannabis consumers’ ability to ride a bicycle” Int J Legal Med DOI 10.1007/s00414-015-1307-y
http://www.ncbi.nlm.nih.gov/pubmed/26739323 add to dropbox
repetitive practical cycling tests and medical examinations were carried out before and after inhalative consumption of cannabis. A maximum of three joints with body weight -adapted THC con-ent (300 μ g THC per kg body weight) could be consumed by each test subject. Fourteen regular cannabis-consuming test subjects were studied (12 males, 2 fe-males). In summary, only a few driving faults were observed even under the influence of very high THC concentrations. A defined THC concentration that leads to an inability to ride a bicycle cannot be presented
Cannabis-related e ffects o n simulated driving p er for-mances did regularly not show relevant impairments of motor behavior, and it is assumed that documented deficits are relat-ed to attention or perception [ 5, 6]
THC consumpti on in doses up to 300 μ g per kilogram body weight is considered to cause relevant cognitive and psychomotor impairments comparable to 0.5 g/l BAC [10]. F u r t he rm o r e, T H C c on ce nt ra ti on s o f 7–10 ng/ml THC in se-rum are thought to evoke comparable impairments to 0.5 g/l BAC. THC serum concentrations below 10 ng/ml should not increase the risk of a traffic accident [ 11 ].
regularly consumed amounts of cannabis varied between approximately 1g per week and 1g per day
exclusion criteria were pregnancy, acute illnesses, history of s ub-st an c e ab us e , int ak e o f ps yc ho a ct i ve med i c a tio n, r ar e consumption of cannabis, and positive urine screening for drugs other t ha n cannabis
Dutch medical cannabis ( Cannabis flos: B edrocan, 22 % dronabinol, <1.0 % cannabidiol; supplier: Dutch Ministry of Health, Welfare and Sport, Office of Medicinal Cannabis, P.O. Box 16114, NL-2500 BC The Hague) was imported for the trials (import authorization no. E5304/2014) with allowance from the German Federal Opium Agency
The consumption of the cannabi s cigarettes w as stan-dardized. Each joint c ontained 300 μ gofTHCperkilo-gram of body weight. The test persons were instruct ed to cons ume the joints in the following way: 4-s i nhalat ion, 10-s holdi ng breath, and 15-s exhalation. A m aximum of three joints could be c onsumed
opthamalogical: The average time required t o r ead the 50-wor d t ext l in-early d i d not significan tly differ befo r e and a f ter t he cons umption of cannabi s… The median time needed
to touch the i nvestigator ’s fingertip 10 times also remained constant and was between 10 and 11 s . The median amplitude of fusion dec reas ed immediately after cannabis c onsumption (Fig. 2a, b) a nd increased again wi th fall ing T HC concent rat ion r esp. CIF. No st at isti cal significances were observed. I t s eems that the simulta-neous influence of amphetamine/MDMA hinders the de-crease i n the amplitude of fusion
t h er e i s n o i nc r e as e i n t h e nu m b er o f d em e ri ts after the cannabi s consumption. While the median de-merits in the CIF groups below 10 a nd above 30 is 8 (Fig. 4b), the m edian demerits decreased from 9 to 8 only when the THC concentrations serve a s a basis. There w ere no s ignificant differences between the THC groups (p =0.81)
The study is limited by the small, male-dominated test group, by the fact that four test persons showed positive results for amphetamine/MDMA, and by the fact that only one test person showed a THC concentration below the limit of detection when the study began. However, the age and gender of the test persons are representative of the group of persons who are regularly found driving under the influence of cannabis products
Some test persons w ith THC concentra-ti ons a bove 100 ng/ ml parti al ly rode t hei r b icycles in a way t hat was not more suspicious than the way the test persons r ode their bicycles during the initial r ides
de Havenon A, Chin B, Thomas K, Afra P. 2011. “The secret “Spice”: an undetectable toxic cause of seizure” Neurohospitalist 1(4): 182-186
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3726077/
“Spice” is an umbrella term for a variety of synthetic cannabinoid products whose inhalation has been associated with an increasing number of toxic side effects resulting in emergency department visits. These side effects (including psychosis, tachyarrhythmia, and seizures) are not typically seen with marijuana (Cannabis sativa) use. We report 2 patients with no prior history of neurological disease that experienced their first generalized tonic–clonic seizure after smoking Spice.
Havens J, Gudino O, Biggs E, Diamond U, Weis J, Cloitre M. 2012. “Identification of Trauma Exposure and PTSD in Adolescent Psychiatric Inpatients: An Exploratory Study” J Trauma Stress 25(2): 171-178
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3742006/
large discrepancy between rates of probable PTSD identified through standardized assessment and during the emergency room psychiatric evaluation (28.6% vs. 2.2%). Adolescents with probable PTSD had greater clinical severity and service utilization, an increased likelihood of being diagnosed with bipolar disorder (27.5% vs. 9.2%) and being prescribed antipsychotic medications (47.5% vs. 27.6%)
Only one patient out of 40 (2.5%) who met criteria for probable PTSD received an admission diagnosis of PTSD
Without knowledge about exposure to a traumatic event, it is virtually impossible to evaluate re-experiencing symptoms… symptoms of avoidance of thoughts and reminders
if the youth or their family does not spontaneously disclose exposure to a traumatic event or if the evaluating clinician does not systematically assess exposure to trauma, it will be impossible to obtain information about these central aspects of PTSD
Hayakawa K, Mishima K, Hazekawa M, Sano K, Irie K, Orito K, Egawa T, Kitamura Y, Uchida N, Nishimura R, Egashira N, Iwasaki K, Fujiwara M. 2008. “Cannabidiol potentiates pharmacological effects of Delta(9)-tetrahydrocannabinol via CB(1) receptor-dependent mechanism” Brain Res 1188: 157-64
http://www.ncbi.nlm.nih.gov/pubmed/18021759
CB1 upregulation enhanced by combining CBD and THC
Heim C, Newport DJ, Heit S, Graham YP, Wilcox M, Bonsall R, Miller AH, Nemeroff CB. 2000 “Pituitary-adrenal and autonomic responses to stress in women after sexual and physical abuse in childhood.” JAMA 284(5):592-7
http://www.ncbi.nlm.nih.gov/pubmed/10918705
Women with childhood abuse exhibited increased pituitary-adrenal and autonomic responses to stress compared with controls. effect particularly robust in women with current symptoms of depression and anxiety.
more than 6-fold greater ACTH response to stress
Heim C, Newport J, Bonsall R, Miller A, Nemeroff C. 2001. “Altered pituitary-adrenal axis responses to provocative challenge tests in adult survivors of childhood abuse” Am J Psychiatry 158(4):575-81
http://ajp.psychiatryonline.org/article.aspx?articleid=174699
Abused women without major depressive disorder exhibited greater ACTH responses to CRF administration, whereas abused women with major depressive disorder and depressed women without early life stress demonstrated blunted ACTH responses.
sensitization of the anterior pituitary and counterregulative adaptation of the adrenal cortex in abused women without major depressive disorder. On subsequent stress exposure, women with a history of childhood abuse may hypersecrete CRF, resulting in down-regulation of adenohypophyseal CRF receptors and symptoms of depression and anxiety
The seeming discrepancy between greater ACTH responses to psychosocial stress and blunted ACTH responses to CRF stimulation in abused women with major depression may reflect the involvement of corticolimbic pathways and enhanced hypothalamic releasing activity (CRF and other releasing factors), thus overriding receptor down-regulation.
Henquet C, Krabbendam L, Spauwen J, Kaplan C, Lieb R, Wittchen H-U, van Os J. 2004. “Prospective cohort study of cannabis use, predisposition for psychosis, and psychotic symptoms in young people.” BMJ 330
http://www.bmj.com/content/330/7481/11
cannabis use was much stronger in those with any predisposition for psychosis at baseline (23.8% adjusted difference in risk, 95% confidence interval 7.9 to 39.7, P = 0.003) than in those without (5.6%, 0.4 to 10.8, P = 0.033
Henquet C, Krabbendam L, Spauwen J, Kaplan C, Lieb R, Wittchen H-U, van Os J. 2005. “Prospective cohort study of cannabis use, predisposition for psychosis, and psychotic symptoms in young people” BMJ 330(7481): 11
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC539839/
after adjustment for age, sex, socioeconomic status, urbanicity, childhood trauma, predisposition for psychosis at baseline, and use of other drugs, tobacco, and alcohol, cannabis use at baseline increased the cumulative incidence of psychotic symptoms at follow up four years later (adjusted odds ratio 1.67, 95% confidence interval 1.13 to 2.46). The effect of cannabis use was much stronger in those with any predisposition for psychosis at baseline (23.8% adjusted difference in risk, 95% confidence interval 7.9 to 39.7, P = 0.003) than in those without (5.6%, 0.4 to 10.8, P = 0.033). The risk difference in the “predisposition” group was significantly greater than the risk difference in the “no predisposition” group (test for interaction 18.2%, 1.6 to 34.8, P = 0.032). There was a dose-response relation with increasing frequency of cannabis use
Henquet C, Murray R, Linszen D, van Os J. 2005. “The environment and schizophrenia: the role of cannabis use” Schizophr Bull 31(3): 608-612
http://schizophreniabulletin.oxfordjournals.org/content/31/3/608.long
although a relative risk of around 2.0 is not a very large effect size, cannabis use is extremely prevalent in young people
although effect sizes were reduced, the effect of cannabis persisted after adjustment for factors such as age, sex, social class, ethnic group, family history of psychiatric illness, urbanicity, and use of other drugs
Henquet C, Rosa A, Krabbendam L, Papiol S, Fananas L, Drukker M, Ramaekers J, van OS. 2006. “An Experimental Study of Catechol-O-Methyltransferase Val158Met Moderation of |[Delta]|-9-Tetrahydrocannabinol-Induced Effects on Psychosis and Cognition” Neuropsychopharmacology 31(12): 2748-57
http://www.nature.com/npp/journal/v31/n12/full/1301197a.html
Carriers of the Val allele were most sensitive to |[Delta]|-9-THC-induced psychotic experiences, but this was conditional on prior evidence of psychometric psychosis liability. |[Delta]|-9-THC impacted negatively on cognitive measures. Carriers of the Val allele were also more sensitive to |[Delta]|-9-THC-induced memory and attention impairments compared to carriers of the Met allele. Experimental effects of |[Delta]|-9-THC on cognition and psychosis are moderated by COMT Val158Met genotype, but the effects may in part be conditional on the additional presence of pre-existing psychosis liability. The association between cannabis and psychosis may represent higher order gene-environment and gene-gene interactions
COMT Valine (Val) allele, the relative risk to develop psychotic illness after cannabis exposure was 10.9, whereas in individuals homozygous for the COMT Methionine (Met) allele, this risk was only 1.1
THC impacted on cognition and psychosis outcomes, but there was evidence that this was conditional on other variables
the differential sensitivity to THC associated withCOMT genotype was in part conditional on additional evidence of psychosis liability, as Val carriers with evidence of psychometric psychosis liability experienced more THC-induced transient psychotic symptoms compared to Val carriers without these additional measures of liability
Higher activity of COMT in the prefrontal cortex, associated with the Val allele, may produce a selective decrease in tonic dopamine subcortically, thus initiating an activation of phasic dopamine transmission. The Met allele, associated with low-activity COMT, is thought to decrease phasic and increase tonic dopamine transmission subcortically (Bilderet al, 2004). Phasic dopamine may be important for flexibility of neuronal activation, whereas tonic dopamine may play a critical role in cognitive stability
we did not replicate differences in THC sensitivity between patients and controls
evidence for differential THC sensitivity was found, restricted to subjects with the Val|[sol]|Val genotype
the hypothesis of higher-order interactions is nevertheless necessary to explain the epidemiological observation that only a small minority of those exposed to cannabis develop a severe psychosis outcome (Henquet et al, 2005b)
liability to schizophrenia may in part consist of a developmentally mediated limitation in the capacity to modulate stress-related increased activity of meso-limbic dopamine neurons (Lieberman et al, 1997).COMT genotypes may be relevant in this regard
Henquet C, Di Forti M, Morrison P, Kuepper R, Murray R. 2008. “Gene-environment interplay between cannabis and psychosis” Schizophrenia Bulletin 34(6): 1111-1121
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2632498/ in dropbox
Cannabis use is considered a contributory cause of schizophrenia and psychotic illness. However, only a small proportion of cannabis users develop psychosis.
Evidence suggests that mechanisms of gene-environment interaction are likely to underlie the association between cannabis and psychosis… multiple variations within multiple genes together with other environmental factors (eg, stress) may interact with cannabis to increase the risk of psychosis
In patients with an established psychotic disorder, cannabis use is associated with more and earlier relapses and poorer psychosocial functioning but perhaps surprisingly also with less negative and affective symptoms
it is manifestly obvious that only a small proportion of cannabis users develop psychosis
Herman J, Perry J, van der Kolk B. 1989. “Childhood trauma in borderline personality disorder” Am J Psychiatry 146(4): 490-5
http://www.ncbi.nlm.nih.gov/pubmed/2929750/
Subjects with borderline personality disorder (N = 21) or borderline traits (N = 11) and nonborderline subjects with closely related diagnoses (N = 23)
Significantly more borderline subjects (81%) gave histories of such trauma, including physical abuse (71%), sexual abuse (68%), and witnessing serious domestic violence (62%); abuse histories were less common in those with borderline traits and least common in the subjects with no borderline diagnosis
Herting M, Schwartz D, Mitchell S, Nagel B. 2010. “Delay discounting behavior and white matter microstructure abnormalities in youth with a family history of alcoholism” Alcoholism: Clinical and Experimental Research 34(9): 1590-1602
http://www.ncbi.nlm.nih.gov/pubmed/20586754 in dropbox
lower FA in the left inferior longitudinal fasciculus and the right optic radiation statistically mediated the relationship between FH status and slower RTs on the delay discounting task… FHP youth showed a trend toward greater impulsive decision making, possibly representing an inherent personal characteristic that may facilitate substance use onset and abuse in high-risk youth
Hester R, Nestor L, Garavan H. 2009. “Impaired error awareness and anterior cingulate cortex hypoactivity in chronic cannabis users” Neuropsychopharmacology 34(11): 2450-2458
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2743772/
Sixteen active chronic cannabis users and 16 control participants were administered a Go/No-go response inhibition task during event-related fMRI data collection
Higher depression scores than controls, more use of alcohol, lifetime joints: 11628.8 ± 5993.4, cannabis abstinence 38.0 ± 47.7, cannabis withdrawal 9.3 ± 2.2 — no reference to abstinence status within text, only within Table 1. Text misleadingly refers to “active” use.
Cannabis users’ inhibitory control performance was equivalent to that of the control group, but the former demonstrated a significant deficit in awareness of commission errors. Cannabis users demonstrated a diminished capacity for monitoring their behaviour that was associated with hypoactivity in the ACC and right insula. In addition, increased levels of hypoactivity in both the ACC and right insula regions were significantly correlated with error awareness rates in the cannabis group (but not controls)
Deficits in error detection have also been found to relate to clinical symptoms, including the debilitating symptoms of loss of insight (Lysaker et al, 1998), perseverative behaviour (Frith, 1987) and delusions of alien control in schizophrenia (Frith and Done, 1989), and poor clinical outcomes (e.g., inability to maintain independent living (Seltzer et al, 1997))
diminished error-related anterior cingulate cortex (ACC) activity in a range of clinical conditions, including schizophrenia (Alain et al, 2002; Carter et al, 2001; Turken et al, 2003), major depression (Steele et al, 2007), and drug addiction (Forman et al, 2004; Kaufman et al, 2003)
The hypoactive error-related ACC response observed in the cannabis group is consistent with similar diminished responses in schizophrenia (Alain et al, 2002; Bates et al, 2002; Carter et al, 2001; Kerns et al, 2005; Laurens et al, 2003; Turken et al, 2003), major depression (Steele et al, 2007), ADHD (Liotti et al, 2005; Rubia et al, 2005) and various drug dependent samples (e.g., cocaine (Kaufman et al, 2003), methamphetamine (London et al, 2005), heroin (Forman et al, 2004), alcohol (Ridderinkhof et al, 2002))
Cannabis users showed less deactivation in the left middle frontal gyrus (BA 10) and right putamen during unaware errors when compared to their aware errors, whereas control participants showed no difference in these regions. The opposite pattern was observed in the right cerebellum, with control participants demonstrating significantly less activity (deactivation) during unaware errors when compared to aware, whereas the cannabis group showed no difference. Common to both groups was the differentiation of aware from unaware errors by greater activity in the right insula, bilateral IPL and right middle frontal cortices, as well as significantly less deactivation in the right middle occipital cortex
higher levels of cannabis craving were associated with poorer error awareness rates
Studies with cannabis users have demonstrated a consistent pattern of increased activity in prefrontal and parietal regions
may indeed precede use and constitute a risk factor for prolonged cannabis use
Hickman M, Vickerman P, Macleod J, Lewis G, Zimmit S, Kirkbride J, Jones P. 2009. “If cannabis caused schizophrenia — how many cannabis users may need to be prevented in order to prevent one case of schizophrenia? England and Wales caculations” Addiction 104(11): 1856-61
http://www.ncbi.nlm.nih.gov/pubmed/19832786
NNP for heavy cannabis and schizophrenia ranged from 2800 [90% confidence interval (CI) 2018-4530] in those aged 20-24 years to 4700 (90% CI 3114-8416) in those aged 35-39. In women, mean NNP for heavy cannabis use and schizophrenia ranged from 5470 (90% CI 3640-9839) in those aged 25-29 to 10 870 (90% CI 6786-22 732) in 35-39-year-olds. Equivalent mean NNP for heavy cannabis use and psychosis were lower, from 1360 (90% CI 1007-2124) in men aged 20-24 and 2480 (90% CI 1408-3518) in women aged 16-19. The mean and median number of light cannabis users that would need to be prevented in order to prevent one case of schizophrenia or psychosis per year are four to five times greater than among heavy users.
Hill A, Williams C, Whalley B, Stephens G. 2012. “Phytocannabinoids as novel therapeutic agents in CNS disorders” Pharmacol Ther 133(1): 79-97
http://www.ncbi.nlm.nih.gov/pubmed/21924288/
The Cannabis sativa herb contains over 100 phytocannabinoid (pCB) compounds and has been used for thousands of years for both recreational and medicinal purposes
Δ(9)-tetrahydrocannabinol (Δ(9)-THC) as the primary mediator of the psychoactive, hyperphagic and some of the potentially therapeutic properties of ingested cannabis… research focus has recently widened to include other pCBs including cannabidiol (CBD), cannabigerol (CBG), Δ(9)tetrahydrocannabivarin (Δ(9)-THCV) and cannabidivarin (CBDV)
Disorders that could be targeted include epilepsy, neurodegenerative diseases, affective disorders and the central modulation of feeding behaviour
Hill M, Gorzalka B. 2004. “Enhancement of anxiety-like responsiveness to the cannabinoid CB1 receptor agonist HU-210 following chronic stress” European Journal of Pharmacology 499: 291-295
http://www.ncbi.nlm.nih.gov/pubmed/15381051 in dropbox
Results demonstrated that in unstressed animals [rats], a low dose of HU-210 induced an anxiolytic response, whereas a high dose induced an anxiogenic response. In stressed animals both the low and high doses of HU-210 induced anxiogenic responses… chronic stress enhances either cannabinoid receptor responsivity or one of the interacting systems implicated in emotional states
Hill M, Hillard C, Bambico F, Patel S, Gorzalka B, Gobbi G. 2009. “The therapeutic potential of the endocannabinoid system for the development of a novel class of antidepressants” Trends in Pharmacological Sciences 30(9): 484-493
http://www.sciencedirect.com/science/article/pii/S0165614709001217
Substantial evidence has accumulated implicating a deficit in endocannabinoid in the etiology of depression; accordingly, pharmacological augmentation of endocannabinoid signaling could be a novel target for the pharmacotherapy of depression
facilitation of endocannabinoid neurotransmission evokes both antidepressant and anxiolytic effects
enhancement of endocannabinoid signaling can enhance serotonergic and noradrenergic transmission; increase cellular plasticity and neurotrophin expression within the hippocampus; and dampen activity within the neuroendocrine stress axis
Hill M, Kumar S, Filipski S, Iverson M, Stuhr K, Keith J, Cravatt B, Hillard C, Chattarji S, McEwen B. 2013. “Disruption of fatty acid hydrolase activity prevents the effects of chronic stress on anxiety and amygdalar microstructure.” Molecular Psychiatry 18(10): 1125-35
http://www.ncbi.nlm.nih.gov/pubmed/22776900
Chronic restraint stress in rats increases FAAH and reduces AEA in the amygdala. All stress induced changes in amygdalar structure and function were absent in mice deficient in FAAH.
Hirvonen J, Goodwin R, Li C-T, Terry G, Zoghbi S, Morse C, Pike V, Volkow N, Huestis M, Innis R. 2012. “Reversible and regionally selective downregulation of brain cannabinoid CB1 receptors in chronic daily cannabis smokers”
http://www.nature.com/mp/journal/v17/n6/full/mp201182a.html in dropbox
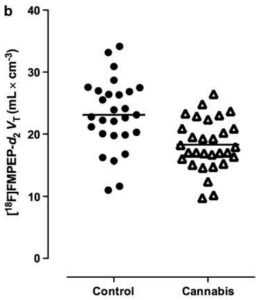
downregulation correlated with years of cannabis smoking and was selective to cortical brain regions. After ~4 weeks of continuously monitored abstinence from cannabis on a secure research unit, CB1 receptor density returned to normal levels
Hohmann A, Suplita R, Bolton N, Neely M, Fegley D, Mangieri R, Krey J, Walker J, Holmes P, Crystal J, Duranti A, Tontini A, Mor M, Tarzia G, Piomelli D. 2005. “An endocannabinoid mechanism for stress-induced analgesia” Nature 435(7045): 1108-12
http://www.ncbi.nlm.nih.gov/pubmed/15973410 in dropbox
Blockade of cannabinoid CB(1) receptors in the periaqueductal grey matter of the midbrain prevents non-opioid stress-induced analgesia. 2-AG and anandamide enzyme blockers (MAGL and FAAH) enhance stress-induced analgesia
stress triggers the formation of both 2-AG and anandamide in the midbrain, but these two endocannabinoids are released with strikingly dissimilar time-courses (2-AG: marked increase 2 min after shock termination, baseline 15 min later; anandamide: peak 7-15 min after shock, persisting but weakening at 25 min)
Holderith N, Nemeth B, Papp O, Veres J, Nagy G, Hajos N. 2011. “Cannabinoids attenuate hippocampal γ oscillations by suppressing excitatory synaptic input onto CA3 pyramidal neurons and fast spiking basket cells” J Physiol 589(20): 4921-34
http://www.ncbi.nlm.nih.gov/pmc/articles/pmid/21859823/
van Holst R, Schilt T. 2011. “Drug-related decrease in neuropsychological functions of abstinent drug users” Curr Drug Abuse Rev 4(1): 42-56
http://www.ncbi.nlm.nih.gov/pubmed/21466500/ – add to dropbox
All substances of abuse, except cannabis, were associated with sustained deficits in executive functioning, especially inhibition. In addition, verbal memory decrements were consistently found in cocaine, (meth)amphetamines and ecstasy users, but not in heroin or cannabis users
There was little evidence for sustained cognitive impairments in adult abstinent cannabis users
a metaanalysis of studies up to 2002 demonstrated no significant detrimental effects of cannabis on neuropsychological functions [75] (Grant)
The short-term effects of cannabis on cognitive functions consist of deficits in attention, working memory, verbal fluency and cognitive flexibility [76]. These deficits are also found in frequent cannabis users after seven days of abstinence [77]… One methodological limitation of these studies is that there was no control for other psychiatric disorders, which could explain some of the differences between cannabis users and controls
although the focus on studies requiring two weeks of abstinence is needed to rule out acute toxification effects, this criterion could also introduce a selection bias. It is conceivable that participants, who are able to refrain from drug use for two weeks or more, actually would like to stop using the substance (as it’s difficult to get non-treatment seekers willing to stop for that long). Moreover, those who are able to remain abstinent might have better executive functions. Conversely, those who want to stop or cut down use might be the people who experience particularly adverse outcomes after drug use. Second, because of the crosssectional nature of most studies, it is impossible to determine whether the identified deficits are a consequence of drug use, or are related to pre-existing vulnerabilities, or a combination of both
Hopper J, Frewen P, van der Kolk B, Lanius R. 2007. “Neural correlates of reexperiencing, avoidance and dissociation in PTSD: symptom dimensions and emotion dysregulation in responses to script-driven trauma imagery” J Trauma Stress 20(5): 713-25
http://www.ncbi.nlm.nih.gov/pubmed/17955540/
reexperiencing severity associated with right anterior insula activity, negatively with right rostral anterior cingulate cortex (rACC). Avoidance correlated negatively with rACC and subcallosal anterior cingulate activity. dissociation correlated positively with activity in the left medial prefrontal and right superior temporal cortices, and negatively with the left superior temporal cortex
Horder J, Cowen P, di Simplicio M, Browning M, Harmer C. 2009. “Acute administration of the cannabionid CB1 antagonist rimonabant impairs positive affective memory in healthy volunteers” Psychopharmacology (Berl) 205(1): 85-91
http://www.ncbi.nlm.nih.gov/pubmed/19337726?dopt=Abstract
A single dose of rimonabant did not alter subjective mood. However, rimonabant selectively reduced incidental recall of positive self-relevant adjectives, an effect contrary to that seen following the administration of antidepressants. There were no effects of rimonabant on the other measures of emotional processing
Horder J, Browning M, di Simplicio M, Cowen P, Harmer C. 2012. “Effects of 7 days of treatment with the cannabinoid type 1 receptor antagonist, rimonabant, on emotional processing” J Psychopharmacol 26(1): 125-32
http://www.ncbi.nlm.nih.gov/pubmed/21406493?dopt=Abstract
Previously, we showed that a single dose of rimonabant produced a negative bias on an emotional word memory task, in the absence of subjective mood effects… In comparison with placebo, rimonabant induced a negative bias on a memory recognition task without producing a change in subjective mood. This raises the possibility that the depressogenic effects of rimonabant may be linked to emotional memory biases
Houston J, Murphy J, Shevlin S, Adamson G. 2011. “Cannabis use and psychosis: revisiting the interactive role of childhood sexual abuse.” Psychol Med 41:2339-2348
http://www.ncbi.nlm.nih.gov/pubmed/21557896 in dropbox
cannabis consumption was predictive of psychosis diagnosis in a bivariate model; within a multivariate model that included childhood sexual abuse, the effect of cannabis use was attenuated and not statistically significant. multivariate analysis revealed non-consensual sex in childhood… over six times [odds ratio (OR) 6.10] more likely to have diagnosis of psychosis compared with those who had not experienced this trauma.”
Howes O, Kapur S. 2009. “The Dopamine Hypothesis of Schizophrenia: Version III — The Final Common Pathway” Schizophr Bull 35(3): 549-562
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2669582/
Huang H, Gundapuneedi T, Rao U. 2012. “White Matter Disruptions in Adolescents Exposed to Childhood Maltreatment and Vulnerability to Psychopathology.” Neuropsychopharmacology 2012: 37, 2693-2701
http://www.ncbi.nlm.nih.gov/pubmed/22850736 in dropbox
adolescents exposed to childhood maltreatment had lower FA values in left and right superior longitudinal fasciculi, right cingulum bundle projecting to the hippocampus, left inferior fronto-occipital fasciculus, and splenium of the corpus callosum compared with controls. observed white matter disruptions SLF-L, SLF-R, and CGH-R associated with vulnerability to unipolar depression and/or substance abuse
effect in right cingulum bundle projecting to hippocampus discriminates between those who went on to develop substance use disorder and those who did not
Hui C, Morcillo C, Russo D, Stochl J, Shelley G, Painter M, Jones P, Perez J. 2013. “Psychiatric morbidity, functioning and quality of life in young paeople at clinical high risk for psychosis” Schizophr Res 148(1-3): 175-180
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3744805/
The presence of co-morbid moderate or severe depressive and anxiety symptoms was common in our sample of young people at enhanced risk for psychosis
Growing evidence is indicating that psychosis may lie on a continuum, with mild psychotic symptoms or psychotic-like experiences at one end and schizophrenia and related psychotic disorders at the other (Kendler et al., 1996; van Os et al., 2001; Dhossche et al., 2002; Johns et al., 2004; van Os et al., 2009). Recent studies including population-based samples also suggest that nearly 80% of the adolescents who report psychotic-like symptoms may have at least one other psychiatric disorder (Kelleher et al., 2012a, 2012b). Furthermore, co-presence of psychotic symptoms in adolescents and young adults with disorders of anxiety and depression appears to be more prevalent than previously considered, and an etiological and functionally relevant feature (Wigman et al., 2012)
60 help-seeking individuals, aged 16–35, referred to CAMEO Early Intervention in Psychosis Service from February 2010 to September 2012 met criteria for HR, according to the Comprehensive Assessment of At Risk Mental States (CAARMS) (Yung et al., 2005). From this assessment, HR individuals were divided into three groups based on whether they were mainly characterised by: i) vulnerability traits (family history of psychosis in first degree relative plus significant drop in functioning levels within past 12 months), ii) attenuated psychotic symptoms, or iii) brief limited intermittent psychotic symptoms (BLIPS). In our sample, all individuals fulfilled criteria for the attenuated psychotic symptoms’ group. Seven individuals (11.7%) also qualified for the vulnerability traits’ group. Intake exclusion criteria included: i) acute intoxication or withdrawal associated with drug or alcohol abuse or any delirium, ii) confirmed intellectual disability (Wechsler Adult Intelligence Scale — tested IQ < 70), or iii) prior total treatment with antipsychotics for more than one week
mean age of 20.7 years (SD = 3.4)… Gender was nearly evenly split between male (n = 55; 52.4%) and female (n = 50; 47.6%)… Less HR individuals achieved higher education degrees (p = 0.001) compared to healthy volunteers, and more HR individuals were single (p = 0.033). A significant proportion of HR individuals were on antidepressant or/and anxiolytic medication (41.7%) at the time of their first contact with CAMEO
MINI DSM-IV diagnoses for 55 of the 60 HR individuals. 38 (69.1%) had more than one DSM-IV psychiatric diagnosis, mainly within the affective and anxiety diagnostic spectra. Primary diagnoses for this group were ranked as follows: major depressive episode, current or recurrent (n = 26; 47.3%) > social phobia (n = 7; 12.7%) = generalised anxiety disorder (n = 7; 12.7%) > obsessive compulsive disorder (n = 5; 9.1%) > bipolar disorder, type II (n = 2; 3.6%) > panic disorder (n = 1; 1.8%) = posttraumatic stress disorder (n = 1; 1.8%). Six HR individuals (10.9%) did not fulfil enough criteria for a DSM-IV Axis I diagnosis
HR individuals had a 72.0% endorsement in suicidal thoughts or intention, as measured with item 9 of BDI-II, whereas only 9.1% of healthy volunteers had positive response in this item (p < 0.001)… 41 HR individuals (85.4%) suffered moderate or severe anxiety symptoms
Approximately 80% of HR individuals had experienced at least one obsessive symptom. Among those who had any obsession or compulsion, the mean of YBOCS total severity score was significantly higher in HR individuals than healthy volunteers (20.1 ± 5.8 vs. 5.3 ± 1.5, p < 0.001), suggesting moderate and subclinical severity respectively
After more than one year of follow-up for each individual at HR in our sample, only 6 (10%) made a transition into FEP. We obtained MINI DSM-IV diagnoses at baseline for 5 of them. 4 had an initial diagnosis of major depression, current or recurrent, and one did not fulfil enough criteria for a DSM-IV mental disorder
HR subjects had poorer functioning, with much lower scores in GAF symptoms and disability than healthy volunteers (45.4 ± 8.9 vs. 86.6 ± 3.8 and 48.6 ± 9.4 vs. 86.7 ± 3.6, respectively, both with p < 0.001), suggesting that individuals with HR mental states suffered serious psychiatric symptoms and any serious impairment in social, occupational or academic functioning. Higher unemployment rate was found in the HR group (37.7% vs. 17.8%, p = 0.029). HR individuals also reported poorer quality of life (3.8 ± 1.0 vs. 5.6 ± 0.6, p < 0.001)
psychotic experiences among young adolescents appear to follow a dose–responseKell pattern in the prediction of a wide variety of future psychopathology (Kelleher et al., 2012a, 2012b)
Our findings highlight the lack of specificity and predictive value of psychotic symptoms and carry important implications for clinicians and researchers in the field of psychosis. Psychotic experiences appear to be common, not only among those patients who suffer from a psychotic illness, but also from other disorders such as depression and anxiety (Wigman et al., 2012)
traumatic events in childhood could eventually manifest as psychotic-like symptoms in the context of non-psychotic psychiatric disorders (Kelleher et al., 2013)
Hungund B, Vinod K, Kassir S, Basavarajappa B, Yalamanchili R, Cooper T, Mann J, Arango V. 2004. “Upregulation of CB1 receptors and agonist-stimulated [35S]GTP S binding in the prefrontal cortex of depressed suicide victims” Molecular Psychiatry 9: 184-190
S binding in the prefrontal cortex of depressed suicide victims” Molecular Psychiatry 9: 184-190
http://www.nature.com/mp/journal/v9/n2/full/4001376a.html
Upregulation of CB1receptor density (Bmax) of 24% observed in brains of depressed suicides compared with matched controls. 38% increase in CB1R immunoreactivity. 45% greater CB1 receptor stimulated protein binding in dorsolateral prefrontal cortex
Decreased CB1 receptor binding in neurodegenerative diseases related to extrapyramidal function has been reported.17 Alterations in the serotonergic (for a review see, Arango et al18) and adrenergic19 receptors in the pathophysiology of depression and suicidal behavior are well documented. Recently, Dean et al,20reported increased CB1 receptor density in dorsolateral prefrontal cortex (DLPFC) in schizophrenia.
The average density (Bmax) of CB1 receptor in DLPFC of normal control subjects was 493.3 52.7 fmol/mg protein
52.7 fmol/mg protein
opiates signaling was found to be 20-fold more efficient than cannabinoid signaling.32 CB1 receptor signaling functions as a subtle, fine-tuning mechanism for cells; high density of receptors makes the CB1 receptor highly sensitive to agonists; however, the poor coupling efficiency ensures that overactivation of the system will not occur
Dowlatshahi et al,33 found decreased cAMP signaling in the brain of depressed suicides; sensitization of cannabinoidergic signaling may lead to the decreased cAMP content of the cell as these receptors are negatively coupled to AC
Aged rats exhibited a marked decrease in CB1receptors and its mediated [35S]GTP S binding sites in rat brain.34 we also observed reduced receptor density with increasing age in normal control subject 15,16
S binding sites in rat brain.34 we also observed reduced receptor density with increasing age in normal control subject 15,16
only three patients had medications or alcohol at the time of death and no psychoactive drugs were detected in the remaining patients
CB1 receptor activation suppresses neurotransmitter release by inhibiting a calcium-dependent step in vesicle release,41 decreasing the local release of synaptic vesicles42,43,44
The pharmacological manipulation of endocannabinoid system may serve as a new therapeutic target in the treatment of depression
Huizink A, Ferdinand R, Ormel J, Verhulst F. 2006. “Hypothalamic-pituitary-adrenal axis activity and early onset of cannabis use” Addiction 101(11):1581-8
http://www.ncbi.nlm.nih.gov/pubmed/17034437
The early onset group had lower cortisol levels 30 minutes after awakening than the late onset group (OR = 0.93, 95% CI: 0.86-0.99). Furthermore, compared to non-users, the early and late onset cannabis users had higher levels of cortisol at 8 p.m
Hutchison K, Haughey H, Niculescu M, Schacht J, Kaiser A, Stitzel J, Horton W, Filbey F. 2008. “The Incentive Salience of Alcohol, Translating the Effects of Genetic Variant in CNR1” Arch Gen Psychiatry 65(7): 841-850
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2856651/
Hyman S, Sinha R. 2009. “Stress-Related Factors in Cannabis Use and Misuse: Implications for Prevention and Treatment” J Subst Abuse Treat 36(4): 400-413
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2696937/
cannabis is commonly used as a stress-coping strategy. Negative life events, trauma, and maladaptive coping related to consumption. Cannabis use for stress-coping purposes was most evident when examining chronic as compared with experimental use
72% of daily cannabis users reported smoking to relax or relieve tension, 36% to escape from problems, 29% to alleviate anger or frustration, 28% to get through the day Johnston and O’Malley (1986)
Green, Kavanagh, and Young (2004): those with psychosis using cannabis to positively alter their mood (36% and 42%), to cope with negative affect (27% and 29%), and for social activity reasons (38% and 29%). Those without using cannabis to relax (34% and 43%), and for social activity reasons (49% and 51%).
stressful life circumstances, especially those that occur early in life, may elevate distress, increase sensitivity to stress, and set individuals on a trajectory toward experimental cannabis use and a more severe course of addiction … cannabis use may be most likely in stress-vulnerable individuals as it supplies immediate stress-relief with relatively little effort
Iannetti G, Wise R. 2007. “BOLD functional MRI in disease and pharmacological studies: room for improvement?” Magnetic Resonance Imaging 25: 978:988
http://www.ncbi.nlm.nih.gov/pubmed/17499469 in dropbox
BOLD fMRI does not measure neural activity directly, but relies on a cascade of physiological events linking neural activity to the generation of MRI signal. However, most of the disease and pharmacological studies performed so far have interpreted changes in BOLD fMRI as bbrain activation,Q ignoring the potential confounds that can arise through drug- or disease-induced modulation of events downstream of the neural activity
necessary improvements consist of (1) the inclusion of one or more control tasks, and (2) the recording of physiological parameters during scanning and subsequent correction of possible between-group differences. Second-line,highly recommended important aim to make the results of a patient or drug BOLD study more interpretable and include the assessment of (1) baseline brain perfusion, (2) vascular reactivity, (3) the inclusion of stimulus-related perfusion fMRI and (4) the recording of electrophysiological responses to the stimulus of interest. Finally, third-line, desirable improvements consist of the inclusion of (1) simultaneous EEG–fMRI, (2) cerebral blood volume and (3) rate of metabolic oxygen consumption measurements and, when relevant, (4) animal studies investigating signalling between neural cells and blood vessels
Insel, T. 2010. “Rethinking schizophrenia” Nature 486: 187-193
http://www.nature.com/nature/journal/v468/n7321/full/nature09552.html in dropbox
A century ago we had large public institutions for serious mental illness, tuberculosis and leprosy. Of these three, today only mental illness, especially schizophrenia, remains unchanged in prevalence and disability
those with serious mental illness were three times more likely to be found in the criminal justice system than in hospitals
Although both conventional and atypical antipsychotics reliably reduce delusions and hallucinations, they have not enhanced functional recovery (for example, employment) for people with schizophrenia. One explanation is that the disability of schizophrenia is largely due to cognitive deficits, such as problems with attention and working memory, which these drugs fail to improve
Although schizophrenic psychosis usually emerges between ages 18–25, several longitudinal population-based studies indicate that problems are evident much earlier. For instance, a recent report from a 45-year follow up of a Copenhagen birth cohort demonstrated that adults with schizophrenia have a history of delayed maturation including delayed developmental milestones in the first year 40 . Data from the Dunedin birth cohort, consist-ent with many previous studies 41, indicated that IQ is reduced early and persistently in children destined to develop schizophrenia 42 These precur-sors of schizophrenia are subtle and non-specific, but the consistency of the finding supports the hypothesis that psychosis does not emerge from a completely healthy brain
Maternal malnutrition during famine 50,51 , infections in the second trimester52 , perinatal injury53 and cytokine exposure 54 have all been associated with subsequent increased risk for schizophrenia
291 adolescents followed for 2.5 years reported that the prodrome represented a 405-fold increase in risk (rela-tive to the general population) and that a combination of three factors (for example, genetic risk with recent functional decline, unusual thought content, and either suspicion/paranoia or reduced social func-tioning) resulted in a positive predictive power for conversion to psychosis of 74–81%
Results from single-site trials of atypical antipsychotics 81 , antidepressants 82, lithium83 and cognitive behaviour therapy 84 have had, at best, modest effects in reducing symptoms or preventing conversion to psychosis
It is now clear that negative symptoms (loss of will, anhedonia, poverty of thought) and cognitive deficits (reduced working memory, poor cognitive control) are core features of the disorder that account for much of the long-term morbidity and poor functional outcomes 87
In 2010, smoking and obesity are epidemic among people with schizophrenia, with estimates of nicotine dependence ranging from 58–90% (ref. 95) and metabolic syndrome (obesity, hyperlipidemia, hyperglycemia and hypertension) present in 40% (ref. 96)
Institute of Medicine. 2009. “Preventing Mental, Emotional and Behavioral Disorders Among Young People: Progress and Possibilities” The National Academies Press
http://www.nap.edu/openbook.php?record_id=12480&page=R1
cost related to ELA ~$247 billion
Ito Y, Teicher M, Glod C, Harper D, Magnus E, Gelbard H. 1993. “Increased prevalence of electrophysiological abnormalities in children with psychological, physical and sexual abuse” Journal of Neuropsychiatry 5:401-408
http://www.ncbi.nlm.nih.gov/pubmed/8286938 in dropbox
27 nonabused, 22 psychologically abused, 55 physical/sexual abuse (within which 38 “unequivocal abuse of substantial severity”)
Abnormal EEG in 26.9% of nonabused, 54.4% of early trauma, 42.9% with psychological abuse, 59.6% in total physical/sexual abuse sample, 71.9% in subsample with definite serious abuse
Majority of abnormalities in frontal, temporal and anterior regions. 2.7 to 3.1 fold greater prevalence of EEG abnormalities localized to frontotemporal and anterior regions than not abused. Mostly left. In non-abused, left abnormalities rare; in abused, left 2.5 more likely abnormal than right. Psychological abuse possibly specific to left temporal. In physical/sexual abuse, increae in prevalence of frontotemporal, and less evidence of left-localisation.
Left-hemisphere deficit indexed by substantially better visual-spatial ability than verbal ability
Recall of traumatic childhood memory shifts cortical activity from left-sided to right-sided; degree of shift correlates with severity of abuse
Irish L, Kobayashi I, Delahanty D. 2009. “Long-term Physical Health Consequences of Childhood Sexual Abuse: A Meta-Analytic Review” Journal of Pediatric Psychology 35(5): 450-461
http://jpepsy.oxfordjournals.org/content/35/5/450.full
53% of patients with functional (non-organic) GI disorders have a history of CSA compared with 37% of those with organic disorders (Drossman,1995).
pain, cardiopulumonary symptoms, obesity, gynecologic health, GI health, general health lack of regular exercise
although abused girls increased body mass at a steeper rate than non-abused girls, the groups did not significantly differ in BMI until early adulthood
differences larger in clinical samples than in community samples
Ishiguro H, Horiuchi Y, Ishikawa M, Koga M, Imai K, Suzuki Y, Morikawa M, Inada T, Watanabe Y, Takahashi M, Someya T, Ujike H, Iwata N, Ozaki N, Onaivi ES, Kunugi H, Sasaki T, Itokawa M, Arai M, Niizato K, Iritani S, Naka I, Ohashi J, Kakita A, Takahashi H, Nawa H, Arinami T. 2010. “Brain cannabinoid CB2 receptor in schizophrenia” Biol Psychiatry 67(10): 974-82
http://www.ncbi.nlm.nih.gov/pubmed/19931854
increased risk of schizophrenia for people with low CB2 receptor function
R63 allele of rs2501432 (R63Q) (p = .001), the C allele of rs12744386 (p = .005) and the haplotype of the R63-C allele (p = 5 x 10(-6)) were significantly increased among 1920 patients with schizophrenia compared with 1920 control subjects in the combined population. A significantly lower response to CB2 ligands in cultured CHO cells transfected with the R63 allele compared with those with Q63, and significantly lower CB2 receptor mRNA and protein levels found in human brain with the CC and CT genotypes of rs12744386 compared with TT genotype were observed. AM630 exacerbated MK-801- or methamphetamine-induced disturbance of prepulse inhibition and hyperactivity in C57BL/6JJmsSlc mice
Jackowski A, Douglas-Palumberi H, Jackowski M, Win L, Schultz R, Staib L, Krystal J, Kaufman J. 2008. “Corpus callosum in maltreated children with PTSD: A diffusion tensor imaging study” Psychiatry Res 162(3): 256-261
http://www.ncbi.nlm.nih.gov/pubmed/18296031 in dropbox
the maltreated children had reduced FA in the medial and posterior regions of the CC
Jackowski A, Perera T, Abdallah C, Garrido G, tang C, Martinez J, Mathew S, Gorman J, Rosenblum L, Smith E, Dwork A, Shungu D, Kaffman A, Gelernter J, Coplan J, Kaufman J. 2011. “Early-life stress, corpus callosum development, hippocampal volumetrics, and anxious behavior in male nonhuman primates” Psychiatry Res 192(1): 37-44
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4090111/
Male bonnet monkeys (Macaca radiata) were subjected to the Variable Foraging Demand (VFD) early stress paradigm as infants
VFD rearing was associated with smaller CC size, CC measurements were found to correlate with fearful behavior in adulthood, and ex-vivo CC assessments showed high consistency with earlier MRI measures. Region of Interest (ROI) hippocampus and whole brain voxel- based morphometry assessments were also completed and VFD rearing was associated with reduced hippocampus and inferior and middle temporal gyri volumes
smaller mid-sagittal cross-sectional area of corpus callosum observed in VFD-reared animals (F1,17=5.43, P <0.032; VFD: 90.0 mm2 ± 11.9; non-VFD: 98.4 mm2 ± 14.6) VBM of total CC volume found no significant main effect
smaller left but not right hippocampal volume in VFD (F1,17=8.21, P < 0.01; VFD: 0.43 cm3 ± 0.04; non-VFD: 0.46 cm3 ± 0.02)
animals were exposed briefly to a masked intruder and fearful responses were scored: emotional responsivity measure correlated significantly with the ROI CC assessments (R=-0.74, P < 0.01) larger mid-sagittal CC = lower intruder stress
Over 80% (5/6) of the VFD subjects with one or two “s” alleles had smaller CC cross-sections… no genotype effect observed for left or right hippocampal volume
Primates who carried at least one “s” allele had smaller volumes of cuneus bilaterally, left fusiform gyrus, and right intraparietal sulcus. Moreover, greater volumes of right amygdala, right striatum, bilateral middle temporal gyrus, right superior temporal gyrus, and right inferior frontal gyrus volumes were observed in the primates with one or two “s” alleles
gender differences in the CC (Bachmann et al., 2003; Suganthy et al., 2003) and the gender-by-early stress effect on the CC (Coe et al., 2002; see )
medial and posterior portions of the CC contain interhemispheric projections from the auditory cortices, posterior cingulate, insula, and somatosensory and visual cortices to a lesser extent. It also includes connections from the inferior parietal lobe to the contralateral inferior parietal lobe, superior temporal sulcus, cingulate, retrosplenial cortex, and parahippocampal gyrus (Pandya and Seltzer, 1986). Several of the regions with interhemispheric projections through the medial and posterior CC have connections with prefrontal cortical areas, and are involved in circuits that mediate the processing of emotional stimuli and various memory functions — core disturbances associated with a history of early trauma
In rodents, early-life stress seems to affect the hippocampus though the effects are seen only in adulthood and not during development (Andersen and Teicher, 2004)
Human studies which showed an association between childhood adverse events and small hippocampal size in adults (Bremner et al., 1997; Stein et al., 1997; Driessen et al., 2000; Vythilingam et al., 2002) but not in children (De Bellis et al., 1999; Carrion et al., 2001; De Bellis et al., 2001; De Bellis, 2002; Tupler and De Bellis, 2006; Woon and Hedges, 2008)
Preclinical studies of the effects of stress suggest a minimum of four mechanisms by which hippocampal atrophy may result from stress: dendritic atrophy, synaptic remodeling (synaptic overproduction and pruning), increased neuronal death, and decreased rates of neurogenesis (Sapolsky et al., 1985; Watanabe et al., 1992; Gould and Cameron, 1996; Sapolsky, 1996; Andersen and Teicher, 2004)
Reductions in middle and inferior temporal gyri were also noted in association with VFD rearing. The middle temporal gyrus and inferior temporal gyrus subserve language and semantic memory processing, visual perception, and multimodal sensory integration (Onitsuka et al., 2004)
Jacobus J, McQueeny T, Bava S, Schweinsburg B, Frank L, Yang T, Tapert S. 2009. “White matter integrity in adolescents with histories of marijuana use and binge drinking” Neurotoxicol Teratol 31(6): 349-355
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2762024/
42 adolescents (ages 16−19) classified as controls, binge drinkers, or binge drinkers who are also heavy marijuana users
Significant between group differences were found in FA in eight white matter regions (ps ≤ .016) between the binge drink-only group and controls, including superior corona radiata, inferior longitudinal fasciculus, inferior fronto-occipital fasciculus, and superior longitudinal fasciculus. Interestingly, in 4 of these same regions, binge drinkers who are also heavy marijuana users had higher FA than binge drinkers who did not use marijuana (ps < .05). MD did not differ between groups
Jacobus J, Thayer R, Trim R, Bava S, Frank L, Tapert S. 2013. “White matter integrity, substance use, and risk taking in adolescence” Psychol Addict Behav 27(2): 431-442
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3416962/
Altered white matter integrity in mid-adolescence was hypothesized to predict subsequent risk taking behaviors 1.5 years later
Adolescent substance users (predominantly alcohol and marijuana, n=47) and demographically similar non-users (n=49)
In substance using youth (n=47), lower white matter integrity at baseline in the fornix and superior corona radiata predicted follow-up substance use (ΔR2 =10–12%, ps < .01), and baseline fornix integrity predicted follow-up delinquent behaviors (ΔR2 = 10%, p < .01) 1.5 years later. Poorer fronto-limbic white matter integrity was linked to a greater propensity for future risk taking behaviors among youth who initiated heavy substance use by mid-adolescence. Most notable were relationships between projection and limbic system fibers and future substance use frequency
While internalizing behaviors have been described as internal emotional states that are “overcontrolled,” externalizing is suggested as “undercontrol,” (Achenbach & Rescorla, 2001; Achenbach & Edelbrock, 1978)
the tendency to engage in some degree of risk taking and novelty seeking can increase opportunities for success and social status (Spear, 2010)
cross-sectional study suggests a link between more coherent white matter in superior/anterior corona radiata fiber tracts and increased engagement in substance use-related risk taking behaviors (Berns, Moore, & Capra, 2009); however this finding is in the opposite directions of neurodevelopmental models of adolescent risk taking and therefore warrants further investigation
Jacobus J, Squeglia L, Bava S, Tapert S. 2013. “White matter characterization of adolescent binge drinking with and without co-occurring marijuana use: A 3-year investigation” Psychiatry Res 214(3): 374-381
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3900025/
controls with consistently minimal substance use, n = 16; teens who gradually increase their heavy episodic drinking n = 17, and continuous binge drinkers with heavy marijuana use, n = 21
chronic alcohol use has been consistently associated with abnormalities in white matter volume and integrity (Kril et al., 1997; Pfefferbaum et al., 2000; Pfefferbaum and Sullivan, 2005; Pfefferbaum et al., 2006)
more alcohol use over 1.5 year follow-up was associated with poorer white matter at follow-up (decreased FA), controlling for marijuana use and baseline white matter health (Bava et al., 2013). Together, findings suggest that deleterious effects might be more attributable to binge drinking than marijuana, as preliminary studies suggest that cannabis may have a lesser effect on white matter development during adolescence (Delisi et al., 2006) and may actually serve a neuroprotective role in attenuating the negative effects of heavy alcohol use on white matter integrity during this critical maturation period (Jacobus et al., 2009)
As existing white matter studies from our laboratory and others are largely cross-sectional, it is unclear if differences between substance users and controls predated the onset or escalation of alcohol and marijuana use, or emerged as a result of substance use
we hypothesized that adolescents who engage in both heavy drinking and/or marijuana would demonstrate poorer white matter integrity compared to controls with minimal substance use histories by late adolescence/early adulthood. We found evidence to support this hypothesis in 15 white matter clusters, including association fiber tracts (e.g., superior longitudinal fasciculus, uncinate fasciculus), projection fiber tracts (e.g., regions of the corona radiata), and interhemispheric tracts (e.g., splenium, genu)… most clusters showed significantly decreased FA from baseline to 3-year follow-up
poorer global neurocognitive performance, as measured by a change from 18-month follow-up to 3-year follow-up was related to poorer white matter integrity within a large white matter tract connecting frontal and parietal brain regions (i.e., superior longitudinal fasciculus) at approximately 20 years of age
we did not see substantial differences in white matter integrity between our two user groups, or correlations between neuropsychological performance and substance use severity (e.g., lifetime drinking occasions, marijuana occasions)
at 18-month follow-up, adolescents who engaged in binge drinking demonstrated better global cognitive performance than their marijuana-using counterparts, but this was no longer observed at the 3-year follow-up
We have also seen alterations in macrostructural integrity (e.g., cortical thickness, (Squeglia et al., 2012b) and functional brain activation patterns (Schweinsburg et al., 2011) in teens with similar alcohol use profiles, as well as declines in cognitive status (Squeglia et al., 2009; Mahmood et al., 2010). In a previous investigation, Jacobus et al. (2009), we found that adolescent alcohol users (approximately 17 years old) demonstrated poorer white matter integrity compared to not only matched controls, but to heavy marijuana users with similar levels of alcohol use
it remains surprising that our marijuana users do not show a marked decrease in tissue integrity compared to the binge drinkers given their consistent and heavy co-occurring use
Jacobson S, Kelleher I, Harley M, Murtagh A, Clarke M, Blanchard M, Connolly C, O’Hanlon E, Garavan H, Cannon M. 2010. “Structural and functional brain correlates of subclinical psychotic symptoms in 11-13 year old schoolchildren” Neuroimage 49(2): 1875-85
http://www.ncbi.nlm.nih.gov/pubmed/19770054?access_num=19770054
11 school-going, non-treatment seeking children aged 11-13 who were at symptomatic risk for psychosis (AR) and 14 healthy control children aged 11-12 without subclinical psychotic symptoms (CON)
fMRI results showed reduced activity in the AR group within right frontal and bilateral temporal cortex for response inhibition and reduced activity within the anterior cingulate, insula and middle frontal gyrus for error-related processing (p<.05, corrected). VBM analysis revealed GM increases in the AR group within middle and superior temporal gyri, angular gyrus, orbitofrontal gyrus and GM decrease within the inferior temporal gyrus (p<.05, corrected). DTI analysis identified WM decreases in the AR group along the inferior fronto-occipital fasciculus, cingulum and inferior longitudinal fasciculus (p<.05, corrected)
Jager G, Kahn R, Van Den Brink W, Van Ree J, Ramsey N. 2006. “Long-term effects of frequent cannabis use on working memory and attention: an fMRI study” Psychopharmacology (Berl) 185(3): 358-68
http://link.springer.com/article/10.1007%2Fs00213-005-0298-7 in dropbox
ten frequent cannabis users (after 1 week of abstinence) less hippocampi activation in MJ users, no structural differences
Jager G, Van Hell H, De Win M, Kahn R, Van Den Brink W, Van Ree J, Ramsey N. 2007. “Effects of frequent cannabis use on hippocampal activity during an associative memory task.” Eur Neuropsychopharmacol 17(4):289-97
http://www.europeanneuropsychopharmacology.com/article/S0924-977X(06)00200-8/abstract
Cannabis users and controls performed equally well during the working memory task and the selective attention task, greater activity in the left superior parietal cortex in the cannabis users
Jansen, K. 2004. Ketamine: Dreams and Realities MAPS
https://www.maps.org/images/pdf/books/K-DreamsKJansenMAPS.pdf
Ketamine binds to receptors (called PCP receptors for historical reasons) inside tunnels and causes a blockage. The outer end of the tunnel is attached to a glutamate receptor (called an NMDA receptor) on the cell surface… A natural tunnel blocker will protect the brain from damage while generating a state of being that also holds the flood of overwhelming anxiety at bay. The NDE may be beneficial for the physical brain as well as having the potential for a positive mental outcome. The benefit may be even greater where the forces of the psyche give the person a strong message to “go back” in terms of a mythological drama, and tell them that it is not their time to go. This might be the final expression of the deep drive in the mind to survive
Jaycox L, Ebener P, Damesek L, Becker K. 2004. “Trauma exposure and retention in adolescent substance abuse treatment” J Trauma Stress 17(2): 113-21
http://www.ncbi.nlm.nih.gov/pubmed/15141784
212 adolescents upon admission to long-term residential drug treatment… 71% trauma exposed, 29% of which PTSD+. Trauma-exposed PTSD- left treatment sooner than nonexposed
Jean-Gilles L, Gran B, Constantinescu C. 2010. “Interaction between cytokines, cannabinoids and the nervous system” Immunobiology 215(8): 606-10
http://www.ncbi.nlm.nih.gov/pubmed/20153076/
Cytokines play important roles in neuroinflammation and neurodegeneration… Activation of the cannabinoid system is associated with therapeutic effects that may be mediated by the down-regulation of cytokine expression. Here we review the findings from studies on the reciprocal regulation of the immune and cannabinoid systems, which is based on functional as well as anatomical connections
Jenko K, Hirvonen J, Henter I, Anderson K, Zoghbi S, Hyde T, Deep-Soboslay A, Innis R, Kleinman J. 2012. “Binding of a tritiated inverse agonist to cannabinoid CB1 receptors is increased in patients with schizophrenia” Schizophr Res 141(2-3): 185-188
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3463751/
The specific binding of [3H]MePPEP to CB1 receptors was 20% higher in patients with schizophrenia than in controls
Jia W, Hegde V, Singh N, Sisco D, Grant S, Nagarkatti M, Nagarkatti P. 2006. “Δ9-Tetrahydrocannabinol-Induced Apoptosis in Jurkat Leukemia T Cells Is Regulated by Translocation of Bad to Mitochondria” Mol Cancer Res 4: 549
http://mcr.aacrjournals.org/content/4/8/549.long
Plant-derived cannabinoids, including Δ9-tetrahydrocannabinol (THC), induce apoptosis in leukemic cells, although the precise mechanism remains unclear
THC down-regulated Raf-1/mitogen-activated protein kinase/ERK kinase (MEK)/ERK/RSK pathway leading to translocation of Bad to mitochondria. THC also decreased the phosphorylation of Akt. However, no significant association of Bad translocation with phosphatidylinositol 3-kinase/Akt and protein kinase A signaling pathways was noted when treated cells were examined in relation to phosphorylation status of Bad by Western blot and localization of Bad to mitochondria by confocal analysis. Furthermore, THC treatment decreased the Bad phosphorylation at Ser112 but failed to alter the level of phospho-Bad on site Ser136 that has been reported to be associated with phosphatidylinositol 3-kinase/Akt signal pathway. Jurkat cells expressing a constitutively active MEK construct were found to be resistant to THC-mediated apoptosis and failed to exhibit decreased phospho-Bad on Ser112 as well as Bad translocation to mitochondria. Finally, use of Bad small interfering RNA reduced the expression of Bad in Jurkat cells leading to increased resistance to THC-mediated apoptosis. Together, these data suggested that Raf-1/MEK/ERK/RSK-mediated Bad translocation played a critical role in THC-induced apoptosis in Jurkat cells
John O, Gross J. 2004. “Healthy and unhealthy emotion regulation: personality processes, individual differences, and life span development” J Pers 72(6): 1301-33
http://www.ncbi.nlm.nih.gov/pubmed/15509284/
we review experimental findings showing that reappraisal has a healthier profile of short-term affective, cognitive, and social consequences than suppression. In the second section, we review individual-difference findings, which show that using reappraisal to regulate emotions is associated with healthier patterns of affect, social functioning, and well-being than is using suppression. In the third section, we consider issues in the development of reappraisal and suppression and provide new evidence for a normative shift toward an increasingly healthy emotion regulation profile during adulthood (i.e., increases in the use of reappraisal and decreases in the use of suppression).
Johns A. 2001. “Psychiatric effects of cannabis” British Journal of Psychiatry 178: 116-122
http://bjp.rcpsych.org/content/178/2/116.full
From New Zealand, a sample of 1000 people aged 18-25 were asked to complete a self-administered questionnaire on cannabis use and related problems (Thomas, 1996). Those respondents who admitted using cannabis (38%) were asked about mental health consequences; of these, 22% reported panic attacks or anxiety. Women were twice as likely as men to report these symptoms. Troisi et al (1998) used urine tests on Italian draftees to identify 133 men who used only cannabis. All individuals with a pre-existing psychosis or severe personality disorder had been excluded. An adjustment disorder with depressed mood was found in 16%, major depression in 14%, and dysthymia in 10.5%. The severity of these symptoms was dose-related. No acute psychotic symptoms were reported. Reilly et al (1998) describe the adverse effects found among 268 cannabis users who had taken the drug for at least 10 years, and who continued to smoke about two refers a day. The most common adverse effects were feelings of anxiety, paranoia or depression (21%), tiredness and low motivation (21%)
Peralta & Cuesta (1992) reported that cannabis had no significant effect on positive symptoms of schizophrenia, but it did attenuate negative symptoms.
Subjects were asked about episodes of violence in the previous year (i.e. hitting a partner, bruising a child, fighting, using a weapon in a fight while drinking). Of the 191 respondents with cannabis abuse or dependence, 19.25% (risk ratio 9.4) had been violent compared with 12.69% (risk ratio 6.2) of those with schizophrenia or schizophreniform disorder and 24.57% (risk ratio 11.9) of those with alcohol abuse or dependence. Here, the risk is expressed relative to the 2.05% who were violent among those of the sample population who showed no psychiatric disorder
Andreasson et al (1987) of 45 570 Swedish conscripts, of whom 9.4% had used cannabis and 1.7% were ‘high consumers’ having used more than 50 times. Fifteen-year follow-up data were drawn from national registers of deaths and psychiatric cases. Compared with non-users, the relative risk of schizophrenia was 2.4 in the group that reported use of cannabis at least once, rising to 6.0 among heavy users. Nearly half (430/730) of these high consumers had a psychiatric diagnosis other than psychosis on conscription; controlling for this reduced the relative risk to 2.9. The authors suggest that cannabis consumption is a ‘ life-event stressor’ for individuals vulnerable to schizophrenia. Hall et al (1994) offer a number of alternative explanations. There is a large temporal gap between self-reported cannabis use on conscription and the development of schizophrenia over 15 years, and no data as to whether the cannabis use continued during this time. Drugs other than cannabis could have been taken at any time after conscription.
It should also be noted that as only 49 of the 274 conscripts with schizophrenia had ever tried cannabis, then this drug may only be relevant to a minority of cases. Furthermore, Jablensky et al (1992) demonstrate a striking uniformity in the incidence of schizophrenia in cultures with very different rates of cannabis consumption.
McGuire et al (1994). In this study, 23 patients with psychosis and with cannabis in their urine were gender-matched with 46 drug-free controls with psychosis, and the lifetime risk of psychiatric disorder among all the first-degree relatives was ascertained. The cannabis-positive subjects had a significantly greater (7.1%) familial risk of schizophrenia than controls (0.7%), suggesting that the development or recurrence of acute psychosis in the context of cannabis use may be associated with a genetic predisposition to schizophrenia.
Johns L, Cannon M, Singleton N, Murray R, Farrell M, Brugha T, Bebbington P, Jenkins R, Meltzer H. 2004. “Prevalence and correlates of self-reported psychotic symptoms in the british population” British Journal of Psychiatry 185(4): 298-305
http://bjp.rcpsych.org/content/185/4/298.long
8580 respondents aged 16–74 years were interviewed
5.5% endorsed one or more items on the PSQ. Factors independently associated with psychotic symptoms were cannabis dependence, alcohol dependence, victimisation, recent stressful life events, lower intellectual ability and neurotic symptoms. Male gender was associated with paranoid thoughts, whereas female gender predicted hallucinatory experiences
n the US National Comorbidity Survey, 28% of individuals endorsed psychosis-screening questions although the rate of clinician-defined psychosis was only 0.7% (Kendler et al, 1996). In the Dunedin birth cohort, 25% of the sample at age 26 years reported at least one delusional or hallucinatory experience that was unrelated to drug use or physical illness, but only 3.7% actually fulfilled criteria for schizophreniform disorder (Poulton et al, 2000). Data on psychotic symptoms were collected as part of the Dutch NEMESIS study (van Os et al, 2000). This representative general population sample of 7076 men and women was interviewed using the Composite International Diagnostic Interview (CIDI), which contains 17 core positive psychosis items; 17.5% of the sample scored at least one on any type of positive psychosis rating but only 2.1% received a DSM–III–R (American Psychiatric Association, 1987) diagnosis of non-affective psychosis. Together, these findings suggest that only a small part of the total phenotypic continuum of psychosis is represented by clinically verified and defined cases… It is likely that variation in prevalence rates across studies is partly a consequence of the different instruments used (number and type of questions asked)
Individuals were 3.5 times as likely (95% CI 2.9–4.5) to experience one or more psychotic symptoms if they scored above 12 on the CIS–R, were almost three times as likely (95% CI 2.0–4.4) if they were dependent on cannabis and were just under 2.5 times as likely (95% CI 1.3–3.9) if they were dependent on any other drug (with or without cannabis). Experience of victimisation and alcohol dependence were also strongly associated with psychotic symptoms (OR=2.0, 95% CI 1.7–2.6; OR=1.8, 95% CI 1.3–2.4, respectively). After controlling for interrelationships between all the variables, young age, non-White ethnic group, urban residence and recent drug use were no longer associated significantly with psychotic symptoms
we cannot determine the direction of the relationship between cannabis dependence and psychotic symptoms from these data… risk was specific to cannabis use, as opposed to the use of other drugs. The association between alcohol dependence and psychotic symptoms may be related to the occurrence of withdrawal symptoms
The association between non-White ethnic group and psychotic symptoms in this study was no longer significant after controlling for other factors, including victimisation and stressful life events. Although it has been reported that Black and ethnic minority patients with psychosis do not experience more life events than do White British patients, they do perceive these events as more threatening (Gilvarry et al, 1999)
experiences of victimisation may lead individuals to believe that they are vulnerable and to view other people and the world as hostile and threatening; and stressful events may then trigger symptoms. Janssen et al (2003) found that perceived discrimination was associated longitudinally with onset of delusional ideation
Johnson K, Bonn-Miller M, Leyro T, Zvolensky M. 2009. “Anxious arousal and anhedonic depression symptoms and the frequency of current marijuana use: testing the mediating role of marijuana-use coping motives among active users” J Stud Alcohol Drugs 70(4): 543-550
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2696295/
After controlling for daily cigarette smoking rate, alcohol consumption, and gender, anxious arousal symptoms, but not anhedonic depression symptoms, were significantly and uniquely associated with the frequency of marijuana use. In addition, coping motives for marijuana use mediated the relation between anxious arousal symptoms and the frequency of current marijuana use.
Jones D, Knosche T, Turner R. 2013. “White matter integrity, fiber count, and other fallacies: the do’s and don’ts of diffusion MRI” 73: 239-54
http://www.ncbi.nlm.nih.gov/pubmed/22846632
Juon H-S, Fothergill K, Green K, Doherty E, Ensminger M. 2011. “Antecedents and consequences of marijuana use trajectories over the life course in an African American population” Drug Alcohol Depend 118(2-3): 216-223
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3161156/
five marijuana trajectories for men (n=455) and four trajectories for women (n=495) were identified extending from adolescence to young adulthood (age 32). We labeled the four trajectory groups similar for men and women “abstainers,” “adolescent only users,” “early adulthood decliners,” and “persistent users.” We named the unique fifth group for men “late starters.”
Although African American youth consistently use marijuana less than white youth (Johnston et al., 2006; Wallace, 1999), the rates among African American adults equal or exceed those of white adults, suggesting a “crossover effect” (Geronimus et al., 1993;National Institute on Drug Abuse, 2002)
In prior work, we have found that first grade children who did not perform well in school were more likely to use drugs later as adolescents (Kellam et al., 1980; Ensminger et al., 1982). Specifically, first grade children rated by teachers as having aggressive behavior or the combination of shy and aggressive behavior were more likely to use alcohol, cigarettes, and marijuana in adolescence and early adulthood (Ensminger et al., 2002). Later in the life course, other social fields, such as family formation and employment, become more prominent, and lack of involvement in these fields relates to later life onset (Green et al., 2010)
ur research is also guided by social control theory (Hirschi, 1969), which explains that those who are strongly bonded to important social institutions are less likely to engage in deviant behavior
Schulenberg et al. (2005) showed that their six marijuana trajectories were related to differences in parental education, religion, high school grades, and employment. Windle and Wiesner (2004), using a white sample, found that their “high chronic” group compared less favorably to the other four trajectory groups on delinquency, academic performance, drug-using friends, and stressful life events
Childhood classroom behaviors were predictors for marijuana trajectories with male later starters(OR=2.13) more likely to be aggressive than abstainers and female adolescent only users (OR=2.51) more likely to be shy than abstainers. Abstainers did not differ on childhood factors from persistent users orearly adult decliners for either men or women
bstainers were more likely to finish high school compared to persistent users(OR=1.88 for men; OR=2.30 for women) and to male early adult decliners (OR=2.60). Female persistent users were more likely to have low adolescent family involvement than abstainers (OR=2.00)
For males in general, those in the three groups with more problematic use (early adult decliners, late starters, and persistent users) were more likely to have a substance use disorder, be incarcerated, never marry, and have high levels of MDD. The male persistent users were at highest risk overall. For females, those in the persistent user group were also at highest risk, with higher levels of substance use disorder, incarceration, and MDD. Interestingly, females in the adolescent only group had the second highest level of MDD, which is intriguing as they would be expected to have fewer mental health problems than those who continue using marijuana into adulthood.
we do not know if the depression influenced trajectory membership, resulted from membership, or merely correlated with the marijuana trajectory
Kaffman A. 2009. “The Silent Epidemic of Neurodevelopmental Injuries” Biol Psychiatry 66(7): 624-626
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2840038/ in dropbox
(licking and grooming in rats protects against hyperactivation of hippocampal corticosterone) because Low LG have lower levels of GR in their hippocampus, they show elevated and prolonged levels of corticosterone when stressed, compared with transient corticosterone release in High LG. once established, this pattern of DNA methylation is highly stable and persists throughout the animal’s life.
Offspring of High LG dams are less fearful, show decreased stress reactivity, and perform better on several hippocampal-dependent tasks compared with offspring raised by Low LG dams
a 2009 report from the Institute of Medicine estimated the cost related to ELA at $247 billion annually (1), which is similar in magnitude to the estimated costs for all cancers combined. Interestingly, however, we spend only $.05 on research related to psychopathology of ELA compared with $2 spent on cancer research for each $100 of their respective costs (12)
Kamprath K, Plendl W, Marsicano G, Deussing J, Wurst W, Lutz B, Wotjak C. 2009. “Endocannabinoids mediate acute fear adaptation via glutamatergic neurons independently of corticotropin-releasing hormone signalling” Genes Brain Behav 8(2): 203-11
http://www.ncbi.nlm.nih.gov/pubmed/19077175/
Kamprath K, Romo-Parra H, Haring M, Gaburro S, Doengi M, Lutz B, Pape H-C. 2011. “Short-term adaptation of conditioned fear responses through endocannabinoid signalling in the central amygdala” Neuropsychopharmacology 36(3): 652-663
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3055679/
CB1 is expressed in the CeA of mice and that CB1 in the CeA mediates short-term synaptic plasticity, namely depolarization-induced suppression of excitation (DSE) and inhibition (DSI). CB1 antagonist AM251 increased both excitatory and inhibitory postsynaptic responses in CeA neurons. Local application of AM251 in the CeA in vivo resulted in an acutely increased fear response in an auditory fear conditioning paradigm. Upon application of AM251 in the basolateral nucleus of the amygdala (BLA) in an otherwise identical protocol, no such acute behavioral effects were detected, but CB1 blockade resulted in increased fear responses during tone exposures on the subsequent days. Moreover, we observed that the efficacy of DSE and DSI in the CeA was increased on the day following fear conditioning, indicating that a single tone-shock pairing resulted in changes in endocannabinoid signaling in the CeA
Kanayama G, Rogowska J, Pope H, Gruber S, Yurgelun-Todd D. 2004. “Spatial working memory in heavy cannabis users: a functional magnetic resonance imaging study” Psychopharmacology (Berl) 176(3-4):239-47
http://www.ncbi.nlm.nih.gov/pubmed/15205869/ in dropbox
12 long-term heavy cannabis users, 6-36 h after last use
Recent cannabis users displayed greater and more widespread brain activation than normal subjects when attempting to perform a spatial working memory task greater dorsolateral prefrontal recruitment
little or no significant correlation with subjects’ years of education, verbal IQ, lifetime episodes of cannabis use, or urinary cannabinoid levels at the time of scanning.
Kaplan H, Tolle G, Yoshida T. 2006. “Substance use-induced diminution of violence: a countervailing effect in longitudinal perspective” Criminology 39(1): 205-224
http://onlinelibrary.wiley.com/doi/10.1111/j.1745-9125.2001.tb00921.x/abstract
cannabis use in adolescence associated with less violence later on
Kaplan B, Springs A, Kaminski N. 2008. “The profile of immune modulation by cannabidiol (CBD) involves deregulation of nuclear factor of activated T cells (NFAT)” Biochem Pharmacol 76(6): 726-737
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2748879/
e demonstrated previously that CBD suppressed interleukin-2 (IL-2) production from phorbol ester plus calcium ionophore (PMA/Io)-activated murine splenocytes. Thus, the focus of the present studies was to further characterize the effect of CBD on immune function. CBD also suppressed IL-2 and interferon-γ (IFN-γ) mRNA expression, proliferation, and cell surface expression of the IL-2 receptor alpha chain, CD25
CBD also suppressed activator protein-1 (AP-1) and nuclear factor of activated T cells (NFAT) transcriptional activity, which are critical regulators of IL-2 and IFN-γ. Furthermore, CBD suppressed the T cell-dependent anti-sheep red blood cell immunoglobulin M antibody forming cell (anti-sRBC IgM AFC) response. Finally, using splenocytes derived from CB1-/-/CB2-/- mice, it was determined that suppression of IL-2 and IFN-γ and suppression of the in vitro anti-sRBC IgM AFC response occurred independently of both CB1 and CB2. However, the magnitude of the immune response to sRBC was significantly depressed in CB1-/-/CB2-/- mice. Taken together, these data suggest that CBD suppresses T cell function and that CB1 and/or CB2 play a critical role in the magnitude of the in vitro anti-sRBC IgM AFC response.
Karagianis J, Novick D, Pecenak J, Haro J, Dossenbach M, Treuer T, Montgomery W, Walton R, Lowry A. 2009. “Worldwide-schizophrenia outpatient health outcomes (W-SOHO): baseline characteristcs of pan-regional observational data from more than 17,000 patients” International Journal of Clinical Practice 63(11): 1578-1588
http://onlinelibrary.wiley.com/doi/10.1111/j.1742-1241.2009.02191.x/abstract
50% (range 41–59%) of patients taking antipsychotics exhibited extrapyramidal symptoms at baseline, and 62% (34–67%) of patients reported sexual dysfunction in the previous month. On average, only 19% (16–23%) of patients were in paid employment and as many as 69% were living in dependent housing
Outcomes were significantly better in “developing” countries than in “developed” countries, possibly due to differences in treatment
Karl A, Schaefer M, Malta L, Dorfel D, Rohleder N, Werner A. 2006. “A meta-analysis of structural brain abnormalities in PTSD” Neurosci Biobehav Rev 30(7): 1004-31
http://www.ncbi.nlm.nih.gov/pubmed/16730374/
smaller hippocampal volumes in PTSD, group differences moderated by MRI methodology, PTSD severity, medication, age and gender. Trauma-exposed PTSD- showed smaller bilateral hippocampal. smaller left amygdala volumes in PTSD compared to both healthy and trauma-exposed controls, and significantly smaller anterior cingulate cortex compared to trauma-exposed controls. Pediatric samples with PTSD exhibited significantly smaller corpus callosum and frontal lobe volumes compared to controls, no group differences in hippocampal volume. (1) hippocampal volumetric differences covary with PTSD severity; (2) hippocampal volumetric differences do not become apparent until adulthood; and (3) PTSD is associated with abnormalities in multiple frontal-limbic system structures
Kathuria S, Gaetani S, Fegley D, Valino F, Duranti A, Tontini A, Mor M, Tarzia G, La Rana G, Calignano A, Giustino A, Tattoli M, Palmery M, Cuomo V, Piomelli D. 2003. “Modulation of anxiety through blockade of anandamide hydrolysis” Nat Med 9(1): 76-81
http://www.ncbi.nlm.nih.gov/pubmed/12461523
in rats FAAH inhibitors exhibit benzodiazepine-like properties in the elevated zero-maze test and suppress isolation-induced vocalizations. These effects are accompanied by augmented brain levels of anandamide and are prevented by CB1 receptor blockade
Kato M, Ohno-Shosaku T, Hashimotodani Y, Uchigashima M, Watanabe M. 2009. “Endocannabinoid-mediated control of synaptic transmission” Physiological Reviews 89(1): 309-380
http://physrev.physiology.org/content/89/1/309.long
Marijuana and other derivatives of the plant Cannabis sativa have been used for thousands of years for their therapeutic and mood-altering properties.
blocking the endocannabinoid system suppresses the extinction of aversive memory (330), relearning of the water maze test (540), cerebellum-dependent eyeblink conditioning (277), drug addiction (323), feeding behavior (407), a certain form of stress-induced analgesia (232), and the recovery of neurobehavioral function after brain injury (411)
CB1 polymorphisms have been reported to link to obesity-related phenotypes (173, 448), hebephrenic schizophrenia (78, 530), childhood attention deficit/hyperactivity disorder (429), and depression in Parkinson’s disease (20)
High levels of [3H]CP55,940 binding are observed in innermost layers of the olfactory bulb, hippocampus (particularly high in the dentate molecular layer and the CA3 region), lateral part of the striatum, target nuclei of the striatum (i.e., globus pallidus, entopeduncular nucleus, substantia nigra pars reticulata), and cerebellar molecular layer. Moderate levels are noted in other forebrain regions and a few nuclei in the brain stem and spinal cord. They include the cerebral cortex (higher in the frontal, parietal, and cingulated areas than other cortical areas), septum, amygdala (nucleus of lateral olfactory tract), hypothalamus (ventromedial hypothalamus), lateral subnucleus of interpeduncular nucleus, parabrachial nucleus, nucleus of solitary tract (caudal and commissural portions), and spinal dorsal horn. The thalamus, other nuclei in the brain stem, and spinal ventral horn are low in ligand binding
CB1 mRNA is expressed in medium spiny neurons and parvalbumin-positive interneurons within the striatum, and in cerebellar granule cells, basket cells, and stellate cells within the cerebellar cortex. In contrast, nonuniform expression reflecting the presence of a few cell types expressing high CB1 mRNA is found in the cerebral cortex, hippocampus, and amygdala. In these regions, strong expression is seen in cholecystokinin (CCK)-positive interneurons, whereas no expression in parvalbumin-positive ineterneurons and generally low expression in principal (or excitatory) neurons are noted
CB1 is preferentially targeted to presynaptic elements (Figs. 1 and 2). As a result, regional distributions of CB1 mRNA and immunoreactivity sometimes dissociate. This is particularly conspicuous when CB1 is predominantly expressed in projection neurons. For example, medium spiny neurons are the output neurons in the striatum, and very intense CB1immunoreactivity is detected in the target regions rather than within the striatum CB1 immunoreactivity is strong along the striatonigral and striatopallidal pathways as well as in substantia nigra pars reticulata and the globus pallidus (Fig. 1A) (342), in both of which CB1mRNA is not expressed
inhibitory synapses generally have higher levels of CB1 than excitatory synapses among CB1-expressing synapses within given neural regions… the density of CB1immunogold labeling on inhibitory synaptic elements is higher than excitatory synapses by 30 times for hippocampal CA1 pyramidal cells (Fig. 2), six times for cerebellar Purkinje cells, and three to four times for striatal medium spiny neurons (267, 528)
CB2 is found in microglial cells, not in astrocytes (13, 387), and is upregulated in response to chronic pain (27, 325, 387, 578)
CB1 activation inhibits adenylyl cyclase or cAMP production in many preparations
In neurons or CB1-transfected cells, application of a cannabinoid agonist activates A-type (198) and inwardly rectifying K+ channels (313) and inhibits N- and P/Q-type Ca2+ channels (524) and D- and M-type K+ channels (365, 466)
neurotransmitters reported to be controlled by the CB1 receptor include glutamate (297), GABA (502), glycine (247), acetylcholine (176), norepinephrine (241), dopamine (61), serotonin (374), and CCK (26)
possible involvement of K+ channels has also been suggested for the suppression of glutamate release at the cerebellar PF-PC synapses (106, 107) and in the NAc (436). Additional involvement of the sites downstream of Ca2+ influx has been demonstrated for the presynaptic suppression of inhibitory (505) and excitatory transmission (570) in the cerebellum
oxytocin is tonically released from oxytocin-producing cells and activates oxytocin receptors, which in turn induces endocannabinoid release and suppresses GABA release through presynaptic CB1 receptors… suggested that oxytocin autoregulates the excitability of magnocellular neurons (307). Endocannabinoid signaling might be involved in this autoregulation by oxytocin.
activation of membrane glucocorticoid receptors induces endocannabinoid release, and thereby suppresses glutamate release through activation of presynaptic CB1 receptors
short-term memory or working memory is highly sensitive, whereas retrieval of previously learned information is resistant to cannabinoids
Systemic administration of the cannabinoid agonist WIN55,212-2 (2.5 and 5 mg/kg) impaired the acquisition of contextual, but not auditory, fear conditioning in rats
administration of WIN55,212-2 facilitated the extinction of contextual fear conditioning at a low dose (0.25 mg/kg), but disrupted it at a higher dose (2.5 mg/kg) in rats
Extinction of aversive memory involves at least two distinct processes, i.e., learning of the association between the conditioned stimulus and the absence of punishment (“associative safety learning”) and habituation to a repeatedly presented stimulus
drugs activating the endocannabinoid system may be useful for the treatments of psychiatric disorders related to retrieval of fear memories, including panic disorders, phobias, and posttraumatic stress disorder (PTSD)
Effects of cannabinoids are dependent on the environmental context, but generally anxiolytic at low doses and anxiogenic at high doses (547)
anxiolytic effects abolished by mu opioid antagonists, anxiogenic effects abolished by k opioid antagonists, but not vice versa.
Activation of CB1 receptors exhibits an antidepressant activity
CB1 antagonists also increased the levels of monoamines including 5-HT, dopamine, and norepinephrine in the prefrontal cortex (525)
Exogenous and endogenous cannabinoids have been shown to exert neuroprotection in a variety of in vitro and in vivo models of neurodegeneration (533
Monory et al. (359) demonstrated that the presence of CB1 receptors in glutamatergic hippocampal neurons is both necessary and sufficient to protect neurons against KA-induced seizures. Therefore, suppression of glutamate release through activation of presynaptic CB1 receptors seems to be a major action of cannabinoids for neuroprotection against KA-induced seizures
Preclinical studies with animal models have suggested the therapeutic potential of cannabinoids for the treatment of neurodegenerative diseases, including multiple sclerosis (103), traumatic brain injury (144, 155), Huntington’s disease (354), and Alzheimer’s disease (354).
its global enhancement as well as global suppression might be of advantage to certain functions, but of disadvantage to others
Katona I, Freund T. 2008. “Endocannabinoid signaling as a synaptic circuit breaker in neurological disease” Nat Med 14(9): 923-30
http://www.ncbi.nlm.nih.gov/pubmed/18776886/
Cannabis sativa is one of the oldest herbal plants in the history of medicine. It was used in various therapeutic applications from pain to epilepsy, but its psychotropic effect has reduced its usage in recent medical practice
endocannabinoid signaling is a key regulator of synaptic communication throughout the central nervous system. Its underlying molecular architecture is highly conserved in synapses from the spinal cord to the neocortex, and as a negative feed-back signal, it provides protection against excess presynaptic activity. The endocannabinoid signaling machinery operates on demand in a synapse-specific manner; therefore, its modulation offers new therapeutic opportunities for the selective control of deleterious neuronal activity in several neurological disorders
Kaufman J, Birmaher B, Perel J, Dahl R, Moreci P, Nelson B, Wells W, Ryan N. 1997. “The corticotropin-releasing hormone challenge in depressed abused, depressed nonabused, and normal control children” Biol Psychiatry 42(8):669-79
http://www.ncbi.nlm.nih.gov/pubmed/9325560
compared to depressed nonabused and normal control children, depressed abused children had significantly greater peak, total, and net ACTH secretion post-CRH… only observed in depressed abused children experiencing ongoing chronic adversity (marital violence, emotional abuse, poverty, lack of supports)
Kearn C, Blake-Palmer K, Daniel E, Mackie K, Glass M. 2005. “Concurrent stimulation of cannabinoid cb1 and dopamine d2 receptors enhances heterodimer formation: a mechanism for receptor cross-talk?” Molecular Pharmacology 67 (5): 1697-1704
http://molpharm.aspetjournals.org/content/67/5/1697?ijkey=8f25b40907343171b591536f982939d23f143f56&keytype2=tf_ipsecsha
Dopamine and endogenous cannabinoids display complex interactions in the basal ganglia. One possible level of interaction is between CB1 cannabinoid and D2 dopamine receptors. Here, we demonstrate that a regulated association of CB1 and D2 receptors profoundly alters CB1 signaling. This provides the first evidence that CB1/D2 receptor complexes exist, are dynamic, and are agonist-regulated with highest complex levels detected when both receptors are stimulated with subsaturating concentrations of agonist. The consequence of this interaction is a differential preference for signaling through a “nonpreferred” G protein. In this case, D2 receptor activation, simultaneously with CB1 receptor stimulation, results in the receptor complex coupling to Gαs protein in preference to the expected Gαi/o proteins. The result of this interaction is an increase in the second messenger cAMP, reversing an initial synergistic inhibition of adenylyl cyclase activity seen at subthreshold concentrations of cannabinoid agonist. Additionally, a pertussis toxin insensitive component in the activation of extracellular signal-regulated kinase (ERK) 1/2 kinases by the cannabinoid agonist CP 55,940 [(1R,3R,4R)-3-[2-hydroxy-4-(1,1-dimethylheptyl)phenyl]-4-(3-hydroxypropyl)cyclohexan-1-ol] is revealed in cells stably expressing both CB1 and D2 receptors. Thus, concurrent receptor stimulation promotes a heterooligomeric receptor complex and an apparent shift of CB1 signaling from a pertussis toxin-sensitive inhibition to a partly pertussis toxin-insensitive stimulation of adenylyl cyclase and ERK 1/2 phosphorylation.
Keiley M, Howe T, Dodge K, Bates J, Pettit G. 2001. “The timing of child physical maltreatment: a cross-domain growth analysis of impact on adolescent externalizing and internalizing problems” Dev Psychopathol 13(4): 891-912
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2769082/
In a sample of 578 children assessed in kindergarten through the eighth grade, we used growth modeling to determine the basic developmental trajectories of mother-reported and teacher-reported externalizing and internalizing behaviors for three physical maltreatment groups of children—early-harmed (prior to age 5 years), later-harmed (age 5 years and over), and nonharmed—controlling for SES and gender. Results demonstrated that the earlier children experienced harsh physical treatment by significant adults, the more likely they were to experience adjustment problems in early adolescence. Over multiple domains, early physical maltreatment was related to more negative sequelae than the same type of maltreatment occurring at later periods
If early emotional bonds are insecure or disorganized, as is often the case for abusive families (Barnett, Ganiban, & Cicchetti, 1999; Carlson, Cicchetti, Barnett, & Braunwald, 1989), and if these bonds are stable, it follows that maltreatment would have more deleterious effects at this time of life as opposed to at a later developmental period when the attachment relationship has already been formed and strengthened
Cicchetti (1989) has suggested that maltreatment may have its greatest impact when self-esteem and self-control processes are developing
Kirby, Chu, and Dill (1993) showed that early age of abuse onset was related to high levels of dissociation in inpatients
no differences in child misbehavior as reported by parents were found between the nonharmed, early harmed, or later harmed children from birth to age 4 years. However, parents of early harmed children (M = .26) reported more child misbehavior at age 4–5 years than did the parents of nonharmed children (M = .19). Parents of later harmed (M = .24) children were not significantly different from the other two sets of parents in reporting child misbehavior between the ages 4 and 5 years
parents of nonharmed (M = .50) children reported using fewer punitive parenting practices (shaking, spanking) at age 4–5 years than did the parents of later harmed (M = .66) children. The parents of early harmed children (M = .55) were not significantly different in punitive parenting practices than either of the other two groups, F (2, 575) =5.28, p < .005
Kelleher I, Harley M, Lynch F, Arseneault L, Fitzpatrick C, Cannon M. 2008. “Associations between childhood trauma, bullying and psychotic symptoms among a school-based adolescent sample” Br J Psychiatry 193(5): 378-82
http://www.ncbi.nlm.nih.gov/pubmed/18978317
Adolescents who reported psychotic symptoms were significantly more likely to have been physically abused in childhood, to have been exposed to domestic violence and to be identified as a bully/victim (that is, both a perpetrator and victim of bullying) than those who did not report such symptoms
Kelleher I, Lynch F, Harley M, Molloy C, Roddy S, Fitzpatrick C, Cannon M. 2012. “Psychotic symptoms in adolescence index risk for suicidal behavior: Findings from 2 population-based case-control clinical interview studies” Arch Gen Psychiatry 69(12): 1277-1283
http://archpsyc.jamanetwork.com/article.aspx?articleid=1386064
Psychotic symptoms were associated with a 10-fold increased odds of any suicidal behavior (ideation, plans, or acts) in both the early and middle adolescence studies (odds ratio [OR], 10.23; 95% CI, 3.25-32.26; P < .001 and OR, 10.5; 95% CI, 3.14-35.17; P < .001, respectively). Adolescents with depressive disorders who also experienced psychotic symptoms were at a nearly 14-fold increased odds of more severe suicidal behavior (suicide plans and suicide acts) compared with adolescents with depressive disorders who did not experience psychotic symptoms (OR, 13.7; 95% CI, 2.1-89.6). Among all adolescents with suicidal ideation, those who also reported psychotic symptoms had a nearly 20-fold increased odds of suicide plans and suicide acts compared with adolescents with suicidal ideation who did not report psychotic symptoms (OR, 19.6; 95% CI, 1.8-216.1)
only 1 of the participants in either of the studies reported command hallucinations to harm or kill themselves
a sense of disintegration and fragmentation of the self resulting from intrusive voices or thoughts have been linked to suicidal thinking.6,37
Jacobson et al42 recently showed volumetric differences in the cingulum and orbitofrontal cortex in a sample of adolescents with psychotic symptoms, 2 centers that are known to play important roles in emotion processing and stress regulation.45 Abnormalities in the orbitofrontal cortex have also recently been highlighted as an area of interest in magnetic resonance imaging studies of suicidal patients.46 Using functional magnetic resonance imaging, we have also demonstrated reduced activity within the right frontal and bilateral temporal cortices during response inhibition tasks in adolescents with psychotic symptoms and, using digital tractography imaging, overall reduced integrity of frontotemporal pathways,42 supporting a profile of a relative disinhibition/proimpulsivity phenotype. These neurobiological findings fit with clinical findings of increased symptoms of emotional and behavioral problems among adolescents with psychotic symptoms.22,27,47
child mental health clinics, where psychosis can sometimes be seen as an “adult psychiatry” issue and therefore not fully explored
Kelleher I, Keeley H, Corcoran P, Lynch F, Fitzpatrick C, Devlin N, Molloy C, Roddy S, Clarke M, Harley M, Arseneault L, Wasserman C, Carli V, Sarchiapone M, Hoven C, Wasserman D, Cannon M. 2012. “Clinicopathological significance of psychotic experiences in non-psychotic young people: evidence from four population-based studies” British Journal of Psychiatry 201)1): 26-32
http://bjp.rcpsych.org/content/201/1/26.long
Younger adolescents had a higher prevalence (21–23%) of psychotic symptoms than older adolescents (7%). In both age groups the majority of adolescents who reported psychotic symptoms had at least one diagnosable non-psychotic psychiatric disorder, although associations with psychopathology increased with age: nearly 80% of the mid-adolescence sample who reported psychotic symptoms had at least one diagnosis, compared with 57% of the early adolescence sample. Adolescents who reported psychotic symptoms were at particularly high risk of having multiple co-occurring diagnoses
Alemany et al have recently published the first study documenting a molecular genetic association, showing an interaction between the BDNF val66met polymorphism and childhood trauma in increasing risk of psychotic symptoms.32
A limitation is that the issue of temporality cannot be resolved in the current study: which came first, psychotic symptoms or psychopathology, or indeed did both arise together?
these findings highlight psychotic symptoms as a complex novel aspect in the study of the aetiology of severe mental illness, of which schizophrenia is only one. Research has only recently begun on neurogenetics, imaging, electrophysiology and cognition among individuals with psychotic symptoms.32,33,42–45
Kelleher I, Keeley H, Corcoran P, Ramsay H, Wasserman C, Carli V, Sarchiapone M, Hoven C, Wasserman D, Cannon M. 2013. “Childhood trauma and psychosis in a prospective cohort study: cause, effect, and directionality” Am J Psychiatry 170(7): 734-41
http://ajp.psychiatryonline.org/doi/full/10.1176/appi.ajp.2012.12091169
Exposure to physical abuse and bullying during childhood was strongly predictive of psychotic experiences in individuals who had never before had symptoms, and stopping the trauma led to a significant drop in the frequency of psychotic experiences
nationally representative prospective cohort study of 1,112 school-based adolescents 13–16 years of age, assessed at baseline and at 3-month and 12-month follow-ups for childhood trauma (physical assault and bullying) and psychotic experiences
A bidirectional relationship was observed between childhood trauma and psychosis, with trauma predicting psychotic experiences over time and vice versa… A dose-response relationship was observed between severity of bullying and risk for psychotic experiences
A total of 111 individuals reported a 12-month history of physical assault at baseline (odds ratio for psychotic experiences, 4.02, 95% CI=2.30–7.05, p<0.001). Of these individuals, 37% also reported at least one physical assault between baseline and the 3-month follow-up. Individuals whose experience of physical assault did not reoccur between baseline and the 3-month follow-up had a lower risk of psychotic symptoms at 3 months compared with those whose experience of physical assault continued (odds ratio=0.17, 95% CI=0.04–0.71, p=0.015) (Figure 1). Similarly, individuals whose experience of physical assault did not reoccur between the 3-month and 12-month follow-ups had a lower risk of psychotic symptoms at 12 months compared with those whose experience of physical assault continued (odds ratio=0.16, 95% CI=0.05–0.53, p<0.003)
Psychotic experiences reported at baseline predicted physical assault at 3 months and at 12 months, even after adjusting for baseline reports of physical assault (3-month odds ratio=7.21, 95% CI=2.30–22.58, p=0.001; 12-month odds ratio=12.56, 95% CI=3.26–48.44, p<0.001). Similarly, psychotic experiences reported at baseline predicted bullying at 3 months, even after adjusting for baseline reports of bullying (3-month odds ratio=5.32, 95% CI=1.16–24.40, p=0.032). There was a nonsignificant trend for psychotic experiences reported at baseline to predict bullying at 12 months after adjusting for baseline reports of bullying (odds ratio=3.22, 95% CI=0.53–19.60, p=0.20
One is that psychotic experiences may lead to behavior that attracts hostile or bullying responses. Another is that children with psychotic experiences may live in social circumstances where traumatic experiences are more likely to happen, in which case psychotic experiences may predict traumatic experiences but not necessarily cause them. A third is that children with psychotic experiences may interpret negative interactions with others in a more paranoid way and may be more likely than children without psychotic experiences to label such experiences as bullying. There is less room for misinterpretation of intention, however, when it comes to physical assault
We would argue that training on the provision of a safe and positive environment that promotes optimal overall child development should form the core of teacher training programs. Formal training on evidence-based interventions to prevent and stop bullying (39) should form the backbone of such a program
Kemeny M. 2009. “Psychobiological responses to social threat: evolution of a psychological model in psychoneuroimmunology” Brain Behav Immun 23(1): 1-9
http://www.ncbi.nlm.nih.gov/pubmed/18809488/
There exists a bidirectional network of interactions between the central nervous system, the endocrine system and the immune system. The existence of these pathways allows stressful life experience to impact the immune system with important implications for health. One powerful elicitor of changes in the autonomic, endocrine and immune systems is threat to social status. This review describes the development of a human model of social status threat that specifies a set of contextual, psychological and biological pathways that may underlie the health consequences of threats to social status and regard. The role of cognitive processes in shaping the physiological response to the social world will be emphasized.
Kempker J, Honig E, Martin G. 2014. “Effects of marijuana exposure on expiratory airflow: a study of adults who participated in the US National Health and Nutrition Examination Study” Ann Am Thorac Soc [epub ahead of print]
http://www.ncbi.nlm.nih.gov/pubmed/25521349
Conclusions: In a large cross-section of U.S. adults, lifetime marijuana use up to 20 joint-years is not associated with adverse changes in spirometric measures of lung health. While > 20 joint-years of marijuana exposure was associated with a two-fold increased odds of a FEV1/FVC < 70%, this was the result of an increase in FVC rather than the disproportional decrease in FEV1 seen with obstructive lung diseases
Kendler K, Bulik C, Silberg J, Hettema J, Myers J, Prescott C. 2000. “Childhood sexual abuse and adult psychiatric and substance use disorders in women: an epidemiological and cotwin control analysis” Arch Gen Psychiatry 57(10): 953-9
http://archpsyc.jamanetwork.com/article.aspx?articleid=481660
1411 female adult twins 30.4% reported any CSA and 8.4% reported intercourse (21% fondling). OR from intercourse: major depression 3.14, generalised anxiety 2.94, panic disorder 2.55, bulimia 3.3, alcohol 4.01, drug dependence 5.7
Khoury L, Tang Y, Bradley B, Cubells J, Ressler K. 2010. “Substance use, childhood traumatic experience, and Posttraumatic Stress Disorder in an urban civilian population” Depress Anxiety 27(12): 1077-1086
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3051362/
587 participants included in this study, all recruited from medical and OB/GYN clinic waiting rooms at Grady Memorial Hospital in Atlanta, GA
high rates of lifetime dependence on various substances were found (39% alcohol, 34.1% cocaine, 6.2% heroin/opiates, and 44.8% marijuana)
Breslau et al., in particular, found that exposure to traumatic experience did not increase the risk of substance problems independently of PTSD symptomology
In all subjects, physical abuse correlated with the use of all substances examined, while sexual abuse in childhood associated with cocaine and marijuana use only
Examination of the childhood trauma quartiles shows alcohol, cocaine, and marijuana use significantly increasing across the four quartiles. This essentially indicates a progressive effect of trauma load on the severity of use of these particular substances… these childhood trauma quartiles represent the number of types of childhood abuse experienced; other important factors may include severity and frequency of abuse, age of first occurrence, as well as perpetrator identity
Although we predicted that this effect of multiple traumatization would not be independent of PTSD symptoms, alcohol and marijuana (but not cocaine) use still increased significantly across childhood trauma quartiles even after controlling for PTSD… we looked at lifetime substance exposure but current PTSD symptoms
childhood trauma contributed to increased alcohol and marijuana use independently of adult trauma exposure
Kim J, Cho B, Cho D, Park H. 2013. “Expression of hypothalamic-pituitary-adrenal axis in common skin diseases: evidence of its association with stress-related disease activity” Acta Derm Venereol 93: 387-393
http://www.ncbi.nlm.nih.gov/pubmed/23462974 in dropbox
CRH is also involved in activating the sympathetic nervous system by stimulating the locus coeruleus of the brain stem
Chronic exposure to stress enhances the central tone of the HPA axis and interferes with the normal function of the HPA axis…. response of which normalizes over time
One hypothesis is that a chronically activated HPA axis could diminish the anti-inflammatory effects of glucocorticoids at the target tissue, which may result from exhausted glucocorticoid levels or tissue receptor resistance
Some patients with chronic inflammatory skin disorders showed a hypo-responsive central HPA axis to acute stress and aberrant peripheral HPA axis hormonal expression in involved skin lesions compared with controls. A few studies have indicated that CRH potentiates the effects of main pathogenic inflammatory cells and their mediators… there may be a feedback loop between central and peripheral HPA axis hormones
Kim M, Chey J, Chung A, Bae S, Khang H, Ham B, Yoon S, Jeong D, Lyoo I. 2008. “Diminished rostral anterior cingulate activity in response to threat-related events in posttraumatic stress disorder” J Psychiatr Res 42(4): 268-77
http://www.ncbi.nlm.nih.gov/pubmed/17400251/
Previous brain imaging studies have reported hyperactivation of the amygdala and hypoactivation of the anterior cingulate in posttraumatic stress disorder (PTSD) patients, which is believed to be an underlying neural mechanism of the PTSD symptoms. The current study specifically focuses on the abnormal activity of the rostral anterior cingulate… found decreased rostral anterior cingulate activation to emotional stimuli
Kim S, Won S, Mao X, Jin K, Greenberg D. 2006. “Molecular mechanisms of cannabinoid protection from neuronal excitotoxicity” Mol Pharmacol 69(3): 691-6
http://www.ncbi.nlm.nih.gov/pubmed/16299067
Cannabinoids seem to protect neurons against NMDA toxicity at least in part by activation of CB1R and downstream inhibition of PKA signaling and NO generation
Kim S, Lee H, Kim H, Bang E, Lee S, Lee D, Woo D, Choi C, Hong K, Lee C, Choe B. 2011. “In vivo and ex vivo evidence for ketamine-induced hyperglutamatergic activity in the cerebral cortex of the rat: potential relevance to schizophrenia” NMR Biomed 24(10): 1235-42
http://www.ncbi.nlm.nih.gov/pubmed/21560175
King J, Mandansky D, King S, Fletcher K, Brewer J. 2001. “Early sexuRaal abuse and low cortisol” Psychiatry Clin Neurosci 55(1): 71-4
http://www.ncbi.nlm.nih.gov/pubmed/11235861
Kishimoto S, Muramatsu M, Gokoh M, Oka S, Waku K, Sugiura T. 2005. “Endogenous cannabinoid receptor ligand induces the migration of human natural killer cells” J Biochem 137(2): 217-23
http://www.ncbi.nlm.nih.gov/pubmed/15749836/
2-arachidonoylglycerol induces the migration of KHYG-1 cells (a natural killer leukemia cell line) and human peripheral blood natural killer cells
anandamide, another endogenous cannabinoid receptor ligand, did not induce the migration. Delta9-tetrahydrocannabinol, a major psychoactive constituent of marijuana, also failed to induce the migration; instead, the addition of delta9-tetrahydrocannabinol together with 2-arachidonoylglycerol abolished the migration induced by 2-arachidonoylglycerol
Kitayama N, Vaccarino V, Kutner M, Weiss P, Bremner J. 2005. “Magnetic resonance imaging (MRI) measurement of hippocampal volume in posttraumatic stress disorder: a meta-analysis” J Affect Disord 88(1): 79-86
http://www.ncbi.nlm.nih.gov/pubmed/16033700
9 studies with 133 adult subjects with chronic PTSD, 148 healthy, and 53 traumatized … significantly smaller volume in both right and left hippocampi in adult subjects with chronic PTSD in comparison with both healthy controls and traumatized controls.
Kitayama N, Quinn S, Bremner J. 2006. “Smaller volume of anterior cingulate cortex in abuse-related posttraumatic stress disorder” J Affect Disord 90(2-3) 171-4
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3226710/
8 subjects with abuse-related PTSD and 13 healthy subjects without PTSD. Right ACC volume in PTSD patients was significantly smaller than in non-PTSD subjects
Kitayama N, Brummer M, Hertz L, Quinn S, Kim Y, Bremner J. 2007. “Morphologic alterations in the corpus callosum in abuse-related posttraumatic stress disorder: a preliminary study” The Journal of nervous and mental disease 195(12): 1027-9
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3229094/
relatively smaller areas of the posterior midbody of the corpus callosum are associated with childhood abuse related PTSD in adults; no reduction in total size or in 7 subregions
9 female subjects with abuse-related PTSD (mean = 37.8 years, SD = 9.8) and 9 female subjects without PTSD and any other psychiatric symptoms
Kessler R, Berglund P, Demler O, Jin R, Merikangas K, Walters E. 2005. “Lifetime Prevalence and Age-of-Onset Distributions of DSM-IV Disorders in the National Comorbidity Survey Replication” Arch Gen Psychiatry 62(6): 593-602
http://archpsyc.jamanetwork.com/article.aspx?articleid=208678
anxiety disorders, 28.8%; mood disorders, 20.8%; impulse-control disorders, 24.8%; substance use disorders, 14.6%; any disorder, 46.4%. Median age of onset is much earlier for anxiety (11 years) and impulse-control (11 years) disorders than for substance use (20 years) and mood (30 years) disorders.
Klein T, Newton C, Larsen K, Lu L, Perkins I, Nong L, Friedman H. 2003. “The cannabinoid system and immune modulation” Journal of Leukocyte Biology 74(4): 486-496
http://www.jleukbio.org/content/74/4/486.long
cannabinoids have been shown to modulate a variety of immune cell functions in humans and animals and more recently, have been shown to modulate T helper cell development, chemotaxis, and tumor development. Many of these drug effects occur through cannabinoid receptor signaling mechanisms and the modulation of cytokines and other gene products. It appears the immunocannabinoid system is involved in regulating the brain-immune axis and might be exploited in future therapies for chronic diseases and immune deficiency
cannabinoids [7 , 8] are also capable of modulating immune function. It is reasonable to speculate that these latter factors serve as secondary modulators, which when mobilized coincident with or shortly after first-line immune modulators such as lymphokines, increase or decrease immune activity
The medicinal uses of marijuana were described centuries ago for diseases such as asthma, migraine, pain, convulsions, and anxiety (reviewed in ref. [9]). More recently, emphasis has been placed on marijuana’s putative, beneficial effects on appetite, glaucoma, spasticity in multiple sclerosis, pain, and inflammation [10]
An influx of Ca2+ activates phospholipases (PL), which drive the conversion of membrane arachidonic acid (AA) to endocannabinoids such as anandamide (ANA), 2-arachidonoylglycerol (2-AG), and 2-arachidonylglyceryl ether (2-AGE). The endocannabinoids can be taken up and metabolized by fatty acid amide hydrolase (FAAH) or can bind to CB1 or CB2. These are G protein-coupled receptors (GPCR) capable of signaling through G protein subunits, Gα, -β, and -γ, leading to a modulation of nitric oxide (NO) and adenylyl cyclase (AC). In neurons, this signaling can lead to a change in K+ and Ca2+ currents and secretion of transmitters such as γ-aminobutyric acid (GABA). In addition, CB1 and CB2 signaling can lead to the activation of many other factors, including PLC, protein kinase C (PKC), nuclear factor-κB (NF-κB), extracellular signal-regulated protein kinase (ERK), focal adhesion kinase (FAK), steroid receptor coactivator (Src), mitogen-activated protein kinase (MAPK), and MAPK kinase (MEK)
ligation of CB1 and CB2 has been shown to not only suppress adenylyl cyclase but also to activate it [50]
moderate marijuana smoking has little in the way of acute effects on systemic immunity but may have significant suppressive effects on the function of immune cells directly exposed to the smoke. For example, marijuana smoking was associated with suppressing the antimicrobial activity, cytokine production, and cytokine responsiveness of pulmonary alveolar macrophages
chronic marijuana smoking increased the CB2 mRNA in PBMC [71]
Stimulation of immune cells also leads to a suppression of CB2 expression
Th cells are powerful regulators of cell-mediated (Th1) and humoral (Th2) adaptive immunity
prostaglandins increase Th2 activity and decrease Th1 activity [2]. In addition, glucocorticoids [98], morphine [99], and certain chemokines [49] are reported to suppress Th1 immunity, and adrenergic agents such as norepinephrine increase Th1 immunity
naïve CD4 Th (Th0) receive immune signals and in addition, receive signals from drugs and endogenous factors in the selective differentiation to Th1 or Th2 cells. Th1 development is decreased (↓) by prostaglandins, glucocorticoids, morphine, chemokines, and cannabinoids and is increased (↑) by norepinephrine. In contrast, Th2 development is decreased (↓) by norepinephrine and increased (↑) by prostaglandins, glucocorticoids, morphine, chemokines, and cannabinoids
THC-injected mice and stimulated in vitro with Con A were shown to produce reduced amounts of IL-2 and IFN-γ
decreased production of IL-12 and increase of IL-10 when pretreated with HU210 and other cannabinoids
In drug-treated cultures, proliferation was inhibited along with Th1 cytokines; Th2 cytokines were increased, and the CB2 antagonist inhibited the effects
Mouse microglia cells also were shown to migrate in response to 2-AG through a CB2-mediated mechanism, and this effect was antagonized by the nonpsychotropic cannabinoids, cannabinol and cannabidiol
cannabinoids are similar to opioids in modulating cell migration and suggest that at least one function of the immunocannabinoid system is to regulate migration of immune cells
Klumpers L, Cole D, Khalili-Mahani N, Soeter R, Rombouts S, van Gerven J. 2012. “Manipulating brain connectivity with δ9 -tetrahydrocannabinol: a pharmacological resting state fMRI study” Neuroimage 63(3): 1701-11
http://www.medicinalgenomics.com/wp-content/uploads/2011/12/FMRI_THC_paper.pdf
Twelve healthy volunteers (9 male, 3 female) inhaled 2, 6 and 6 mg THC or placebo with 90-minute intervals in a randomized, double blind, cross-over trial
THC administration increased functional connectivity in the sensorimotor network, and was associated with dissociable lateralized connectivity changes in the right and left dorsal visual stream networks. The brain regions showing connectivity changes included the cerebellum and dorsal frontal cortical regions
Clear increases were found for feeling high, external perception, heart rate and cortisol, whereas prolactin decreased
THC induces both increases and (to a lesser extent) decreases in functional brain connectivity, mainly in brain regions with high densities of CB1-receptors
Nine subjects dropped out due to adverse effects during THC occasions only [how did they manage to have half of their sample drop out due to adverse effects?]
The bilateral and left dmPFC, and the left frontal pole and left superior frontal gyrus (SFG) had an increased connectivity to the right dorsal visual stream network, whereas the right superior frontal pole, right dorsolateral PFC (dlPFC), and the right inferior and middle frontal gyri had a decreased connectivity
in healthy subjects, THC demonstrated no effects on cognitive functions such as planning and reasoning in the very limited available literature (Morrison et al., 2009; Ramaekers et al., 2009)
the right posterior pole and lateral occipital 580 cortex had an increased connectivity with the left visual dorsal stream 581 network. The occipital regions are functional visual areas (Hine, 1918; 582 Kolmel, 1988). Previous studies found that THC influences several as- 583 pects of vision that could be attributed to visual cortex changes
Most subjects experienced the familiar feelings of subjective ‘high’ after administration of THC. This raises the question of which net- 626 works could be associated with these psychomimetic effects [familiar feeling = psychomimetic?]
our results should be interpreted with the caveat that some of the regional drug effects might be confounded with signal change due to vascular motion
Koenen K, Moffitt T, Caspi A, Taylor A, Purcell S. 2003. “Domestic violence is associated with environmental suppression of IQ in young children” Deve Psychopathol 15(2): 297-311
http://www.ncbi.nlm.nih.gov/pubmed/12931829
eight point 8 drop from environmental exposure to domestic violence in a large scale twin study (n=1116)
Koethe D, Giuffrida A, Schreiber D, Hellmich M, Schultze-Lutter F, Ruhrmann S, Klosterkotter J, Piomelli D, Leweke F. 2011. “Anandamide elevation in cerebrospinal fluid in initial prodromal states of psychosis” Br J Psychiatry 194(4): 371-2
http://www.ncbi.nlm.nih.gov/pubmed/19336792/
Cerebrospinal anandamide levels in patients were significantly elevated. Patients with lower levels showed a higher risk for transiting to psychosis earlier. This anandamidergic up-regulation in the initial prodromal course may suggest a protective role of the endocannabinoid system in early schizophrenia
Koldzic-Zivanovic N, Tu H, Juelich T, Rady P, Tyring S, Hudnall S, Smith E, Hughes T. 2006. “Regulation of adrenal glucocorticoid synthesis by interleukin-10: a preponderance of IL-10 receptor in the adrenal zona fasciculata” Brain Behav Immun 20(5): 460-8
http://www.ncbi.nlm.nih.gov/pubmed/16256304/
To date most cytokines have been shown to be pro-inflammatory, such as interleukin (IL)-1, tumor necrosis factor (TNFalpha), and IL-6
IL-10 inhibits steroid production (StAR) (measured by the production of the corticosterone precursor, progesterone), the expression of steroidogenic acute regulatory protein (semi-quantitative RT-PCR), as well as the activity of the proximal steroidogenic enzymes P450scc and/or 3beta-hydroxysteroid dehydrogenase (3beta-HSD) (measured by progesterone production in 22(R)-hydroxycholesterol-treated cells). Interestingly, all of the above-mentioned effects of IL-10 occur through its inhibition of ACTH effects, but not by IL-10 alone
the region of the adrenal gland responsible for the vast majority of corticosterone synthesis, the zona fasciculata, predominantly expresses the IL-10 receptor 1 (IL-10R1), with little expression in the zona glomerulosa and reticularis. These data demonstrate that IL-10 could play an important role in the regulation of glucocorticoid biosynthesis and in maintenance of homeostasis and immunity during periods of stress
van der Kolk B, Saporta J. 1991. “The biological response to psychic trauma: mechanisms and treatment of intrusion and numbing” Anxiety Research (U.K.) 4: 199-212
http://www.cirp.org/library/psych/vanderkolk2/
The recognition that trauma is qualitatively different from stress and results in lasting biological emergency responses following traumatic experiences may account for the biphasic trauma response, and the accompanying memory disturbances “physioneurosis”.
A century ago, Pierre Janet (1889) taught that overwhelming experiences are accompanied by “vehement emotions” which interfere with proper information processing and appropriate action. He thought that this hyperarousal caused the characteristic memory disturbances that accompany traumatization, by interfering with information processing on a verbal, symbolic level. Hyperarousal causes memories to be split off from consciousness and to be stored as visual images or bodily sensations. Fragments of these “visceral” memories return later as physiological reactions, emotional states, nightmares, flashbacks, or behavioral reenactments (van der Kolk & van der Hart, 1989)
Janet thought that the original excessive physiological response to trauma accounted for the continued emergency responses to subsequent stresses. He claimed that fear needs to be tamed for proper cognitive appraisal and for appropriate action: experiences which overwhelmed people’s coping mechanisms set the stage (or to use Pavlov’s later concept “condition” them) to react automatically with excessive emotional reactions to current experiences rooted in the past.
Freud adopted these views from Janet and also suggested that the fixation on the trauma is biologically based: “After severe shock . . . the dream life continually takes the patient back to the situation of his disaster from which he awakens with renewed terror . . . the patient has undergone a physical fixation to the trauma” (Freud, 1919, 1954, p. 207) The feature of hyperactivity to external stimuli was described by Freud in the clearest neuropsychiatric terms that he knew: “I think that one may venture . . . the traumatic neurosis as the result of an extensive rupture in the barrier against stimuli . . . we seek to understand the effect of the shock by considering the breaking through of the barrier with which the psychic organ is provided (p. 207).
Pavlov’s investigations continued the tradition of explaining the trauma response as the result of lasting physiological alterations (Pavlov, 1926). He, and others employing his paradigm, coined the term “defensive reaction” for a cluster of innate reflexive responses to environmental threat. Many studies have shown how the response to potent environmental stimuli (unconditional stimuli-US) becomes a conditioned reaction. After repeated aversive stimulation, intrinsically non-threatening cues associated with the trauma (conditional stimuli-US) becomes a conditioned reaction. After repeated aversive stimulation, intrinsically non-threatening cues associated with trauma (conditional stimuli-CS) become capable of eliciting the defensive reaction by themselves (conditional response-CR). A rape victim may respond to conditioned stimuli, such as the approach by an unknown man as if she were about to be raped again, and experience panic. Pavlov also pointed out that “constitutional factors”, i.e. individual differences in temperament, accounted for the variability in the human approach to traumatic stimuli.
Abraham Kardiner (1941) who first systematically defined post-traumatic stress for American audiences, noted that sufferers from PTSD continue to live in the emotional environment of the traumatic event, with enduring vigilance for and sensitivity to environmental threat. He described the five principal features of PTSD as (1) persistance of startle response and irritability, (2) proclivity to explosive outbursts of aggression, (3) fixation on the trauma, (4) constriction of the general level of personality functioning, nad (5) atypical dream life. He suggested that the startle reaction probably was a conditioned reflex and considered it the central element of the post-traumatic stress reaction, relating it to the development of irritability and psychosomatic symptoms in these patients.
In Men under Stress, Grinker and Spiegel (1945) describe physical symptoms in the acute post-traumatic state that seem to reflect neurochemical changes of the catecholamine system: they describe flexor changes in posture, hyperkinesis, “violently propulsive gait”, tremor at rest, masklike faces, absence of associated movement while walking, cogwheel rigidity, gastric distress, urinary incontinence, mutism, and a violent startle reflex. Grinker and Spiegal noted the similarity of many of these symptoms and those of diseases of the extrapyramidal motor system. They seem to depict an extraordinary stimulation of biological systems, implicating ascending amine projections in particular
The phasic post-traumatic symptoms of hyperalertness, hyper-reactivity to stimuli and traumatic reexperiencing have been documented in a vast literature on combat trauma, crimes, such as rape (e.g., Burgess & Holstrom, 1974; Kilpatrick, Veronen, & Best, 1985), kidnapping (Terr, 1983), natural disasters (e.g., Shore, Tatum & Vollmer, 1986), accidents (e.g., Wilkinson, 1983) and imprisonment (Krystal, 1978). The human response to trauma is so constant across traumatic stimuli that it is safe to say that the central nervous system (CNS) seems to react to any overwhelming, threatening and uncontrollable experience in quite a consistent pattern
respond to stress in an all-or-nothing way
while people with PTSD tend to deal with their environment by emotional constriction, their bodies continue to react to certain physical and emotional stimuli as if there were a continuing threat of annihilation
Mallow, Fairbank, and Keane (1983) Kolb and Multipassi (1982), Blanchard, Kolb, Geradi, Ryan, and Pallmyer (1986) and Pitman, Orr, Forque, deJong, and Claiborn (1987), all have found significant conditioned reactions in response to stimuli reminscent of the original trauma, as measured by heartrate, bloodpressure and electromyogram. … lack habituation to acoustic startle
Rainey et al. (1987) showed lactate, which stimulates the physiological arousal system, elicited PTSD-like flashbacks in 7/7 subjects and panic attacks in 6 in 7 patients with PTSD, 6 of whom also met panic disorder criteria. Southwick that yohimbine injections (which stimulate NE release from Locus Coeruleus) induce somatosensory flashbacks in people with PTSD (Southwick et al., submitted)
The loss of neuromodulation that is at the core of PTSD leads to intensification of emotional reactivity in general: traumatized people go immediately from stimulus to response without being able to make the intervening psychological assessment of the cause of their arousal, which causes them to overreact and intimidate others. Non-specific noises played into the rooms of sleeping people with post-traumatic stress may precipitate nightmares in which old traumatic occurrences are recreated in exact detail (Kramer, Schoen, & Kinney, 1984). Hyperorousal also interferes with psychotherapy, in preventing remembering and working through painful memories.
Numbing of Responsiveness Numbing of responsiveness, which may be registered as depression, as anhedonia and amotivational states, as psychosomatic reactions, or in dissociative states, is tonic and part of the patients’ baseline functioning. It interferes with the ability to explore, remember and symbolize which are essential to finding good meaning
“the trauma spectrum” (van der Kolk, 1988): other post-traumatic disorders ranging from those that result from brief traumatic exposure at an early age, such as phobias and panic, to Borderline Personality Disorder and Multiple Personality Disorder which are usually associated with chronic intrafamilial abuse (Herman, Perry, and van der Kolk, 1989).
Non-brain damaged adult patients who mutilate themselves invariably seem to have a history of severe childhood trauma, and their behavior has been associated with abnormalities of the endogenous opioid and catecholamine systems (Bach-y-Rita, 1974), van der Kolk, Greenberg, Orr, & Pitman, 1989)
disruption of early attachment directly affects the maturation of the limbic system (Kling & Steklis, 1976)
The hippocampus, which records in memory the spatial and temporal dimensions of experiences, dose not fully mature until the third or fourth year of life. However, the system that subserves memories related to the quality (feel and sound) of experience (which is located in the amygdala) matures much earlier (O’Keefe & Nadel, 1978; Jacobs & Nadel, 1985). Thus, in the first few years of life only the quality of events, but not their context can be remembered
A third trauma related function of the limbic system involves the issue of kindling. Intermittent stimulation of the limbic system with an electrical current that was initially too small to produce overt behavioral effects can eventually sensitize limbic neuronal circuits and lower neuronal firing thresholds: repeated stimulation of the amygdala causes long-term alterations in neuronal excitability (for a review, see van der Kolk, 1987). It is possible that similar kindling phenomena occur when people are repeatedly traumatized, or when one traumatic event is followed by intrusive reexperiences
The function of the septo-hippocampal system is to evaluate the rough meaning of incoming stimuli, and whether they are associated with reward, punishment, novelty, or non-reward. The hippocampus thus is thought to be the evaluation center involved in behavioral inhibition, obsessional thinking, exploratory behavior, scanning and construction of a spatial map (O’Keefe & Nadel, 1978)
External stress increases corticosterone production which decreases the firing rate of the hippocampuse (Pfaff, Silva, & Weiss, 1971). Lesions of the hippocampus lead to motor paralysis because of excessive interference from competing responses
The crucial role of the septo hippocampal system is to activate a descending inhibatory pathway which prevents initiation of emergency responses until it is clear that they are needed
serotonergic antagonists also cause increased aggression in response to stress, and hyper-reactivity to stimuli. (Sheard & Davis, 1976). The suppression of behavior by punishment is reversed by serotonin receptor blockers (e.g., Cook & Sepinwall, 1975). The ascending serotonergic pathways are thought to signal the septo-hippocampus to distinguish punishment from reward
Arousal The body responds to increased physical or psychological demands by releasing norepinephrine from the Locus Coerulus and adrenorcorticotrophin (ACTH) from the anterior pituitary
Peripherally, the body’s stress response consists of the secretion of norepinephrine by the sympathetic nerves and of epinephine by the adrenal medulla, while, stimulated by ACTH, the adrenal cortex secretes glucocorticoids
Perry and associates (Perry, Giller, & Southwick, 1987) have demonstrated a 40% decrease in the muber of platelet alpha2 adrenergic receptors in 25 patients with PTSD. Lerer, Ebstein, Shestatsky, Shemesh, and Greenberg (1987) recently reported evidence of desensitization of adenylate cyclase coupled adrenergic receptors on lymphocytes and platelets with PTSD
serotonergic input into the septal-hippocamus decreases the relative strength of the noradrenergic input, allowing modulation of the emergency responses
severely stressed animals, opiate withdrawal symptoms can be produced equally by termination of the stressful stimulus or by naxolone injections … severe, chronic stress in animals results in a physiological state which resembles dependence on high levels of exogenous opioids (Terman, Shavit, Lewis, Cannon, & Liebeskind, 1984; Maier & Seligman, 1976)
We found that two decades after the original trauma, people with PTSD developed opioid-mediated analgesia in response to a stimulus resembling the traumatic stressor, which we correlated with a secretion of endogenous opioids equivalent to 8 mg. of morphine
triad of physical analgesia, psychic numbing and depersonalization
they may experience physical analgesia, dissociative reactions and emotional numbing, which can be abolished b (1) an act of self-mutilation or (2) the administration of naloxone (Richardson & Zaleski, 1983; Sandman, Barron, Crinella, & Donnelly, 1987)
Since autonomic arousal is mediated by the LC it is plausible that not only flashbacks, but also traumatic nightmares accur following autonomic nervous system activation, mediated by the potentiated pathways from the LC to the hippocampus and amygdala. This also could account for the eidectic (picure-like), rather than oneiric (dream-like) quality of traumatic nightmares (van der Kolk et al., 1984)
PTSD patients to be exquisitely sensitive to non-specific auditory stimuli during sleep; the resulting autonomic arousal seems to precipitate nightmares about traumatic experiences… post-traumatic nightmares occur during any stage of the sleep cycle; that most tend to occur during 2 and 3 a.m. during Stage II or III sleep, possibly during a transition to REM sleep. When they occur during Stage II or III, patient report exact living experiences of traumatic material, while, during REM, they are more likely anxiety dreams
tricyclic antidepressants are effective in treating affective disorders in patients with PTSD, but do no little for core PTSD symptomatology, including psychological numbing
Kolliakou A, Castle D, Sallis H, Joseph C, O’Conner J, Wiffen B, Gayer-Anderson C, McQueen G, Taylor H, Bonaccorso S, Gaughran F, Smith S, Greenwood K, Murray R, Di Forti M, Atakan Z, Ismail K. 2015. “Reasons for cannabis use in first-episode psychosis: Does strength of endorsement change over 12 months?” European Psychiatry 30(1): 152-159
https://kclpure.kcl.ac.uk/portal/en/publications/reasons-for-cannabis-use-in-firstepisode-psychosis(601aa65e-b910-4447-9b49-2cf8dec031ca).html
At each time-point, patients endorsed ‘enhancement’ followed by ‘coping with unpleasant affect’ and ‘social motive’ more highly for their cannabis use than any other reason. ‘Conformity and acceptance’ followed closely. ‘Relief of positive symptoms and side effects’ was the least endorsed motive. Conclusions: Patients endorsed their reasons for use at 3 months and 12 months less strongly than at baseline. Little support for the self-medication or alleviation of dysphoria models was found. Rather, patients rated ‘enhancement’ most highly for their cannabis use
MV: apparently these writers don’t know what “negative symptoms” are or recognise enhancement as relief… astounding ignorance from King’s College London
Konings M, Stefanis N, Kuepper R, de Graaf R, ten Have M, van Os J. 2012. “Replication in two independent population-based samples that childhood maltreatment and cannabis use synergistically impact on psychosis risk” Psychological Medicine 42: 149-159
http://www.ncbi.nlm.nih.gov/pubmed/21676285 in dropbox
the psychosis-inducing effects of cannabis being elevated only in those with the highest level of physical punishment (‘ often ’) in childhood (Table 1). For these individuals, the adjusted effect of cannabis on psychosis outcome was much stronger ( B=1.46, 95 % CI0.87–2.06, p<0.001), compared to those with physical punishment rated ‘ occasionally ’ (B=0.55, 95 % CI 0.30–0.81, p<0.001) or ‘ never ’ (B=0.55, 95 % CI0.11–0.99, p=0.015).
maltreatment moderates the effects of cannabis in a dose-dependent, extra-linear fashion, more severe maltreatment being associated with the greatest effect of cannabis in later expression of psychosis
psychosis at baseline did not predict future cannabis use
Sensitization involving dopaminergic signaling has been proposed as a possible mechanism by which environmental factors such as stress or cannabis use impact on psychosis risk (Collip et al . 2008)
Rats living under normal conditions (i.e. access to water and food), that were exposed to THC, showed only minor behavioral changes and no change in dopaminergic neurotransmission (MacLean & Littleton, 1977). By contrast, under stressful housing conditions (i.e. isolation and food deprivation), THC administration had marked behavioral consequences
cannabis use was dichotomized as ‘ never ’ versus ‘ at least once ’. Lifetime use of other drugs was similarly dichotomized as ‘ never ’ versus ‘at least once ’.
Koola M, McMahon R, Wehring H, Liu F, Mackowick K, Warren K, Feldman S, Shim J-C, Love R, Kelly D. 2012. “Alcohol and cannabis use and mortality in people with schizophrenia and related psychotic disorders” J Psychiatr Res 46(8): 987-993
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3392453/
Out of 762 subjects, 62 died during follow up. In a Cox model, predicted mortality risk was higher in age group 35–55 compared to <35 years and in males, but reduced in cannabis users. Overall five- (3.1% vs 7.5%) and ten-year mortality risk (5.5% vs. 13.6%) was lower in cannabis users than in non-users with psychotic disorders (p=0.005) in a survival model
Koolschijn P, van Haren N, Lensvelt-Mulders G, Hulshoff Pol H, Kahn R. 2009. “Brain volume abnormalities in major depressive disorder: a meta-analysis of magnetic resonance imaging studies” Hum Brain Mapp 30(11): 3719-35
http://www.ncbi.nlm.nih.gov/pubmed/19441021?access_num=19441021&link_type=MED&dopt=Abstract
In this comprehensive meta-analysis the magnitude and extent of brain volume differences between 2,418 patients with major depressive disorder and 1,974 healthy individuals from 64 studies was determined
Patients showed large volume reductions in frontal regions, especially in the anterior cingulate and orbitofrontal cortex with smaller reductions in the prefrontal cortex. The hippocampus, the putamen and caudate nucleus showed moderate volume reductions
Kosten T, Ambrosio E. 2002. “HPA axis function and drug addictive behaviors: insights from studies with Lewis and Fischer 344 inbred rats” Psychoneuroendocrinology 27(1-2): 35-69
http://www.ncbi.nlm.nih.gov/pubmed/11750769
Kucewicz M, Tricklebank M, Bogacz R, Jones M. 2011. “Dysfunctional prefrontal cortical network activity and interactions following cannabinoid receptor activation” Journal of Neuroscience 31(43): 15560-15568
http://www.jneurosci.org/content/31/43/15560.full
CP5540-induced deficits in hippocampal theta and prefrontal gamma oscillations were observed alongside disrupted theta-frequency coherence between the two structures
Kupfer D, Detre T, Koral J, Fajans P. 1973. “A comment on the ‘Amotivational Syndrome’ in marijuana smokers” American Journal of Psychiatry 130(12): 1319-1322
http://ajp.psychiatryonline.org/doi/abs/10.1176/ajp.130.12.1319
Heavy smokers and light smokers of marijuana were compared for general psychological health, stable personality traits, changes in personality induced by long-term use of marijuana, effects of multiple drug use, and the “amotivational syndrome” that is said to result from heavy use of the drug. Results did not indicate any overall pattern of excess psychopathology for either group, although significant differences were found in levels of depression and organicity and in various personality traits. The findings suggest that heavy use of marijuana may be related to already existing depression and that impaired motivation may be a manifestation of depression rather than a consequence of frequent mairjuana use.
Kushner M, Riggs D, Foa E, Miller S. 1993. “Perceived controllability and the development of posttraumatic stress disorder (PTSD) in crime victims” Behav Res Ther 31(1): 105-10
http://www.ncbi.nlm.nih.gov/pubmed/8417720/
three factors; perceived controllability felt during the assault, expected controllability over future assaults, and perceived controllability over aversive events more generally. Only the latter factor was associated with PTSD symptom severity. The hypothesis that perceived controllability would be negatively associated with assault severity was partially supported. Further analyses showed that the association between controllability and PTSD was not mediated or moderated by assault severity measures
Laaris N, Good C, Lupica C. 2010. “Delta9-tetrahydrocannabinol is a full agonist at CB1 receptors on GABA neuron axon terminals in the hippocampus” Neuropharmacology 59(1-2): 121-7
http://www.ncbi.nlm.nih.gov/pubmed/20417220/
concentration-response curves revealed that the maximal effects of Delta(9)-THC and WIN55,212-2 were similar, indicating that Delta(9)-THC is a full agonist at CB1Rs on GABA axon terminals. These results suggest that Delta(9)-THC inhibits GABA release, but does not directly alter GABA(A) receptors or GABA uptake in the hippocampus. Furthermore, full agonist effects of Delta(9)-THC on IPSCs likely result from a much higher expression of CB1Rs on GABA versus glutamate axon terminals in the hippocampus
Lachenmeier D, Rehm J. 2015. “Comparative risk assessment of alcohol, tobacco, cannabis and other illicit drugs using the margin of exposure approach” Sci Rep 5: 8126
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4311234/
The toxicological MOE approach validates epidemiological and social science-based drug ranking approaches especially in regard to the positions of alcohol and tobacco (high risk) and cannabis (low risk)
the results confirm that the risk of cannabis may have been overestimated in the past
Ladarre D, Roland A, Biedzinski S, Ricobaraza A, Lenkei Z. 2014. “Polazrized cellular patterns of endocannabinoid production and detection shape cannabinoid signalling in neurons” Front Cell Neurosci 8: 426
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4285097/
somatodendritic 2-AG levels differently modified signaling responses to CB1R activation by Δ9-THC, the psychoactive compound of marijuana, and by the synthetic cannabinoids WIN55,212-2 and CP55,940. These highly contrasted differences in sub-neuronal signaling responses warrant caution in extrapolating pharmacological profiles, which are typically obtained in non-polarized cells, to predict in vivo responses of axonal (i.e., presynaptic) GPCRs
Polarized neuronal architecture maintains the directionality of information flow through neuronal networks. Accordingly, protein and lipid composition of the plasma membrane greatly differs between axons and the somatodendritic compartment (Horton and Ehlers, 2003). Local interaction between cell membrane components is increasingly considered as a key dynamic component in sensory and signaling pathways. Notably, the highly regulated lipid environment may control the structure, conformation and function of embedded proteins (Phillips et al., 2009). A major brain G-protein coupled receptor (GPCR) that may be particularly sensitive to the lipid composition of the plasma membrane is the type-1 cannabinoid receptor (CB1R)
eCBs are generally considered to be retrograde signals, being produced in the postsynaptic cell and traveling “backwards” across the synaptic cleft to activate CB1Rs on presynaptic nerve terminals (Freund et al., 2003; Kano et al., 2009). However, in addition to this retrograde synaptic signaling effect, eCBs synthetized in the somatodendritic membrane may also have cell-autonomous effects on local CB1Rs, such as endocannabinoid-mediated somatodendritic slow self-inhibition (SSI) (Bacci et al., 2004; Marinelli et al., 2009) or somatodendritic endocytosis-driven transcytotic targeting (Leterrier et al., 2006; Simon et al., 2013)
In axons, the combined effect of high CB1R density and absence of DAGLα activity leads both to elevated response amplitude following agonist stimulation, as well as to a lack of constitutive activation. In the somatodendritic compartment, relatively low CB1R density and high DAGLα activity, locally producing the membrane component endocannabinoid 2-AG, results in constitutive activation of CB1R-activated signaling which is accompanied by significant but relatively low amplitude agonist-induced signaling responses
both WIN and THC require the presence of endogenous 2-AG to efficiently activate CB1Rs.
Lally N, Nugent A, Luckenbaugh D, Ameli R, Roiser J, Zarate C. 2014. “Anti-anhedonic effect of ketamine and its neural correlates in treatment-resistant bipolar depression” Translational Psychiatry 4, e469
http://www.nature.com/tp/journal/v4/n10/full/tp2014105a.html
reduction occurred independently from reductions in general depressive symptoms. Anti-anhedonic effects were specifically related to increased glucose metabolism in the dorsal anterior cingulate cortex and putamen. Our study emphasizes the importance of the glutamatergic system in treatment-refractory bipolar depression, particularly in the treatment of symptoms such as anhedonia
Lane S, Cherek D, Lieving L, Tcheremissine O. 2005. “Marijuana effects on human forgetting functions” J Exp Anal Behav 83(1): 67-83
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1193701/
both Δ9-THC doses impaired delay-dependent discrimination but not delay-independent discrimination
The acute administration of smoked marijuana to adult humans produced changes in rates of forgetting but not in initial discriminability
Based on both pharmacological data and studies of patients with Alzheimer’s disease, White and Ruske (2002) have concluded that disruption of memory related to the cholinergic (i.e., acetylcholine) system is due to impairment in initial discrimination rather than rates of forgetting
Lanius R, Williamson P, Hopper J, Densmore M, Boksman K, Gupta M, Neufeld R, Gati J, Menon R. 2003. “Recall of emotional states in posttraumatic stress disorder: an fMRI investigation” Biol Psychiatry 53(3): 204-10
http://www.ncbi.nlm.nih.gov/pubmed/12559652/
Compared to the trauma-exposed comparison group, PTSD subjects showed significantly less activation of the thalamus and the anterior cingulate gyrus (area 32) in all three emotional states (sad, anxious, and traumatic)
Lee K, Williams L, Haig A, Gordon E. 2003. “”Gamma (40 Hz) phase synchronicity” and symptom dimensions in schizophrenia” Cogn Neuropsychiatry 8(1): 57-71
http://www.ncbi.nlm.nih.gov/pubmed/16571550
Psychomotor Poverty showed decreased left hemisphere synchrony; Reality Distortion was associated with increased right synchrony; Disorganisation showed a widespread enhancement with a delay in frontal Gamma synchrony
widespread and excessive in Disorganisation, localised and enhanced in Reality Distortion, versus localised and diminished in Psychomotor Poverty
Le Strat Y, Le Foll B. 2011. “Obesity and Cannabis Use: Results from 2 Representative National Surveys” American Journal of Epidemiology doi: 10.1093/aje/kwr200
Levita L, Bois C, Healey A, Smyllie E, Papkonstantinou E, Hartley T, Lever C. 2014. “The Behavioural Inhibition System, anxiety and hippocampal volume in a non-clinical population” Biology of Mood & Anxiety Disorders 4(4)
http://www.biolmoodanxietydisord.com/content/4/1/4
Leweke F, Giuffrida A, Wurster U, Emrich H, Piomelli D. 1999. “Elevated endogenous cannabinoids in schizophrenia” Neuroreport 10(8): 1665-9
http://www.ncbi.nlm.nih.gov/pubmed/10501554
erebrospinal concentrations of two endogenous cannabinoids (anandamide and palmitylethanolamide) were significantly higher in schizophrenic patients than non-schizophrenic controls (p < 0.05). By contrast, levels of 2-arachidonylglycerol, another endogenous cannabinoid lipid, were below detection in both groups
Leweke FM, Koethe D. 2008. “Cannabis and psychiatric disorders: it is not only addiction.” 13(2):264-75
Cannabis use is not only frequent in patients suffering from Bipolar Disorder, but it also induces manic symptoms in this group
Li X, Alpati V, Jackson C, Xia S, Bertisch H, Branch C, Delisi L. 2012. “Structural abnormalities in language circuits in genetic high-risk subjects and schizophrenia patients” Psychiatry Res 201(3): 182-9
http://www.ncbi.nlm.nih.gov/pubmed/22512952?access_num=22512952
decrease of the normal left-greater-than-right anatomical asymmetry in the inferior orbital frontal area, and a increased left-greater-than-right pattern in the inferior parietal and occipital regions
Liang M, Zhou Y, Jiang T, Liu Z, Tian L, Liu H, Hao Y. 2005. “Widespread functional disconnectivity in schizophrenia with resting-state functional magnetic resonance imaging” Neuroreport 17(2): 209-213
http://www.ncbi.nlm.nih.gov/pubmed/16407773 in dropbox
the disconnection hypothesis for this disorder has become popular, which assumes that schizo-phrenia could arise from dysfunctional integration of a distributed network of brain regions
(fMRI) and diffusion tensor imaging on schizophrenia has supported the disconnection hypothesis, indicating improper functional integration within the brain
The disconnectivities between many cortical and subcortical regions in schizophrenia, such as the frontal lobe, the temporal lobe, the parietal lobe, the basal ganglia, the thalamus and the cerebellum, have been reported using functional connectivity with fMRI (e.g. [2–4])
we examined the functional con-nectivity of the entire brain of patients with schizophrenia by dividing the entire brain into 116 regions automatically and performing correlation analysis on each pair of these regions
PreF, prefrontal lobe; O thF, o ther par ts of frontal lobe excluding prefrontal lobe; Par, parie tal lobe; Occ , occipital lob e; Tem, temp or al lobe including medial temp or al s truc tures; Ins, insula ; CS, corpus s tria tum; Tha , thala-mus; Cer, cerebellum interestingly, we see that the increased
functional connectivities in schizophrenia are mainly related to the cerebellum
In our study, schizophrenic patients mainly showed decreased functional connectivities (158 out of a total of 177 abnormal functional connectivities) during the resting state compared with controls. This result supports the hypothesis of disconnection in schizophrenia.
Boksman et al. [3] reported a widespread unfocussed interaction between the right anterior cingulate and other parts of the brain in schizophrenic patients, while normal controls showed a
localized interaction between the right anterior cingulate and the left temporal lobe using a word fluency paradigm. In addition, some diffusion tensor imaging studies have focused on the anatomical disconnection of schizophrenia and reported reduced fractional anisotropy of the whole-brain white matter, the frontotemporal lobe, the corpus callosum, the anterior cingulum and various other brain regions [5]
Relative to other areas, four regions including the insula, the temporal lobe (including the medial temporal struc-tures), the prefrontal lobe and the corpus striatum exhibited a larger number of decreased functional connectivities after normalization in patients (Fig. 1c). Many studies have focused on dysfunctional prefrontal, temporal lobe and corpus striatum and the connectivities related to them and have suggested that these were very important in the pathophysiology of schizophrenia [6,19–21]. Although there were fewer studies relating to the involvement of the insula in schizophrenia, several of these have indicated an abnormal volume and function of the insula in schizophrenia [22–24]. Our results showed decreased functional connectivities between the insula and these three regions, the prefrontal lobe, the temporal lobe and the corpus striatum, during rest
Lightman S. 2008. “The neuroendocrinology of stress: a never ending story” J Neuroendocrinol 20(6): 880-4
http://www.ncbi.nlm.nih.gov/pubmed/18601712/
hormones (i.e. corticosterone in the rat and cortisol in man) are released in a pulsatile ultradian pattern which defines the normal circadian rhythm. The frequency of the pulses is increased under states of chronic stress, and in rats with genetically determined hyper-responsiveness of the hypothalamic-pituitary-adrenal axis. Interestingly, neonatal influences can also programme alterations in ultradian rhythmicity, implicating epigenetic factors in its regulation. At the level of tissue receptors, the alteration in pattern of glucocorticoid ultradian rhythm has differential effects on mineralocorticoid receptor and glucocorticoid receptor (GR) binding to DNA and offers a mechanism for tissue specific responses to altered glucocorticoid dynamics. The effects of neonatal experience are not only seen at the level of CRH and GR regulation, but also are evident in behavioural responses to stress and in the responsiveness of brain stem serotonergic pathways, as measured by tryptophan hydroxylase mRNA in the brain stem
Linszen D, Dingemans P, Lenior M. 1994. “Cannabis abuse and the course of recent-onset schizophrenic disorders” Arch Gen Psychiatry 51(4): 273-279
http://archpsyc.jamanetwork.com/article.aspx?articleid=496554
Cannabis-abusing patients (n=24) were compared with nonabusers (n=69). Eleven patients were mild and 13 were heavy cannabis-abusing patients
Significantly more and earlier psychotic relapses occurred in the cannabis-abusing group (P=.03). This association became stronger when mild and heavy cannabis abuse were distinguished
Lipschitz D, Rasmusson A, Anyan W, Gueorguieva R, Billingslea E, Cromwell P, Southwick S. 2003. “Posttraumatic stress disorder and substance use in inner-city adolescent girls” J Nerv Ment Dis 191(11): 714-21
http://www.ncbi.nlm.nih.gov/pubmed/14614338
Compared with traumatized girls without PTSD, girls with full and partial PTSD were significantly more likely to use nicotine, marijuana, and/or alcohol on a regular basis. Fifteen girls met criteria for both PTSD and a substance-use disorder. For 80% of these girls, the age of onset of PTSD was either before or concurrent with the onset of their substance-use disorder
Lisdahl K, Wright N, Medina-Kirchner C, Maple K, Shollenbarger S. 2014. “Considering cannabis: the effects of regular cannabis use on neurocognition in adolescents and young adults” Curr Addict Rep 1: 144-156
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4084860/
During manuscript preparation, Dr. Lisdahl was supported by the NIDA-funded grant R01DA030354
Due to Krista Lisdahl’s efforts, the University of Wisconsin-Milwaukee has received money in the form of grants from NIH/NIDA for the current work. Dr. Lisdahl declares consultancy work for the NIH/NIDA and travel, accommodations or meeting expenses paid by INS, APA and NIDA, outside of the submitted work
data is not clear as to whether the age limit should be set at 21, or closer to 25 when significant gray matter neurodevelopment is complete
several cross-sectional studies in cannabis-using youth without psychiatric comorbidities report cannabis-related cognitive deficits including reduced processing speed (Fried, Watkinson, & Gray, 2005; Gruber et al., 2012; Lisdahl & Price, 2012; Medina, Hanson, et al., 2007), complex attention (Hanson, et al., 2010; Harvey, Sellman, Porter, & Frampton, 2007; Lisdahl & Price, 2012; Mathias et al., 2011; Medina, Hanson et al., 2007; Tapert et al., 2002), verbal memory (Fried et al., 2005; Hanson, et al., 2010; Harvey et al., 2007; McHale & Hunt, 2008; Medina, Hanson, et al., 2007;Schwartz, Gruenewald, Klitzner, & Fedio, 1989; Solowij et al., 2011; Tait et al., 2011; Thoma et al., 2011), executive functioning (Gonzalez et al., 2012; Grant et al., 2012; Gruber et al., 2012; Hanson et al., 2010;Harvey et al., 2007; Lisdahl & Price, 2012; Mathias et al., 2011; McHale & Hunt, 2008; Medina, Hanson et al., 2007; Schuster et al., 2012; Solowij et al., 2012), and risky sexual behavior (Schuster et al., 2012)
we found that female cannabis users had larger posterior PFC (Medina et al., 2009), posterior inferior cerebellar vermis (Medina et al., 2010), and left amygdala (McQueeny et al., 2011) volumes. Male users also had larger posterior inferior cerebellar vermis volumes (Medina et al., 2010). Churchwell and colleagues (2012) found increased striatal volume in a sample of comorbid cannabis and methamphetamine users. Other groups have reported decreased right medial orbitofrontal cortex (Churchwell et al., 2010), reduced hippocampal (Ashtari et al., 2011; Cousijn et al., 2012; Schacht et al., 2012), reduced amygdala (Cousijn et al., 2012; Schacht et al., 2012) and increased anterior cerebellar (Cousijn et al., 2012) volumes in adolescent and young adult cannabis users without comorbid psychiatric conditions
Lopez-Larson and colleagues (2011) measured cortical thickness in cannabis-users and found decreased cortical thickness in right caudal middle frontal, bilateral insula, and bilateral superior frontal cortices with increased thickness in lingual, temporal, inferior parietal, and paracentral areas in the cannabis users compared to non-users
magnetic resonance spectroscopy research has shown alterations in glutamate, N-acetyl aspartate, creatine, and myo-inositol in the anterior cingulate (Prescot et al., 2011), reduced subcortical global myo-inositol/creatine ratios, and reduced white matter myo-inositol (Silveri et al., 2011)
chronic cannabis use during the adolescent and emerging adult years may result in abnormal vascular functioning as well as neuronal and migroglia toxicity. Additional studies are needed to determine whether vascular and glial alterations underlie GM and WM abnormalities observed in young cannabis users
(fMRI) and positron emission tomography (PET) studies that assess brain activation patterns in adolescent cannabis users have reported abnormal PFC, parietal, insular, subcortical/limbic, and cerebellar activation in cannabis users during finger tapping (Lopez-Larson et al., 2012), attentional control (Abdullaev et al., 2010), spatial working memory (Schweinsburg et al., 2008;Schweinsburg et al., 2010; Smith et al., 2010), verbal working memory (Jacobsen, Pugh, Constable, Westerveld, & Mencl, 2007; Jager, Block, Luijten, & Ramsey, 2010), verbal learning (Becker et al., 2010), executive functioning (Tapert et al., 2007; Vaidya et al., 2012), and pleasant interoceptive stimuli (Migliorini et al., 2013) tasks. In the latter study, findings demonstrated that substance-using teens had blunted processing of pleasant stimuli but heighted sensitivity to reward processing in the insula (Migliorini et al., 2013)
Increased parietal-cerebellar, right hemispheric, and PFC (Behan et al., 2013; Orr et al., 2013) and decreased frontal-cerebellar interhemispheric and temporal cortex (Houck et al., 2013; Orr et al., 2013) resting state connectivity has been reported in adolescent cannabis users. Studies have also found increased functional connectivity between PFC-occipital (Harding et al., 2012), and parietal-cerebellar (Behan et al., 2013) brain regions in adolescent cannabis users while engaging in cognitive tasks
Without additional large-scale prospective longitudinal studies, it can be difficult to tease apart the influence of premorbid factors versus direct effects of cannabis exposure on neurocognition in youth. Most of the studies outlined in this review controlled for potentially confounding variables, such as family history of SUD and Axis I disorders, however, subclinical symptoms or neurocognitive correlates of risk factors (Clark et al., 2012; Hanson, et al., 2010; Hill, Muddasani et al., 2007; Nigg et al., 2004;Schweinsburg et al., 2004; Spadoni, et al., 2008; Ridenour et al., 2009; Tapert & Brown, 2000) may still explain at least a portion of the brain abnormalities reported in cross-sectional studies of cannabis users
Conclusions: increase prevention and treatment
most of the studies examined youth following a period of abstinence and excluded individuals with comorbid disorders; therefore, these findings may underestimate functioning of young cannabis users who may be experiencing withdrawal symptoms, poor sleep quality, comorbid psychiatric symptoms, and stress associated with increased legal issues
It needs to be emphasized that regular cannabis use, defined here as once a week, is not safe and may result in addiction and neurocognitive damage, especially in youth
increase resources for prevention, screening, and early intervention for regular cannabis users (especially targeting youth)
Liu H, Liu Z, Liang M, Hao Y, Tan L, Kuang F, Yi Y, Xu L, Jiang T. 2005. “Decreased regional homogeneity in schizophrenia: a resting state functional magnetic resonance imaging study” NeuroReport 17(1): 19-22
http://www.ncbi.nlm.nih.gov/pubmed/16361943 in dropbox
decreased regional homogeneity in schizophrenia, which distributed over the bilateral frontal, temporal, occipital, cerebellar posterior, right parietal and left limbic lobes
reduced frontal blood flow/metabolism in resting state schizophrenia
López H. 2010. “Cannabinoid-hormone interactions in the regulation of motivational processes” Hormones and Behavior 58: 100-110
http://www.ncbi.nlm.nih.gov/pubmed/19819241 in dropbox
There is a bi-directionality in hormone-cannabinoid interactions: cannabinoids affect prominent endocrine axes (such as the hypothalamic-pituitary-gonadal), and gonadal hormones modulate cannabinoid effects.
Lopez-Quintero C, Hasin D, de los Cobos J, Pines A, Wang S, Grant B, Blanco C. 2011. “Probability and predictors of remission from lifetime nicotine, alcohol, cannabis or cocaine dependence: results from the National Epidemiologic Survey on Alcohol and Related Conditions” Addiction 106(3): 657-669
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3227547/
Lifetime probability of dependence remission nicotine 83.7%; alcohol 90.6%; cannabis 97.2%; cocaine 99.2%. Half by 26, 14, 6 and 5 years after onset. Personality disorder, blacks and males have lower rate of remission
43,093, representative of US population. Dependence n’s: nicotine 6,937, alcohol 4,781, cannabis 530, cocaine 408.
Cannabis dependence comorbidity: any 98.95%, mood 56.95%, anxiety 56.07%, conduct 2.74%, personality 65.38%, nicotine 69.64%, alcohol 72.15%, cocaine 23.14%, family substance use disorder 70.75%, onset before age 14 35.35%
Lorenzetti V, Lubman D, Whittle S, Solowij N, Yücel M. 2010. “Structural MRI findings in long-term cannabis users: what do we know?” Subst Use Misuse 45(11):1787-808
http://www.ncbi.nlm.nih.gov/pubmed/20590400
No study found global structural changes in cannabis users, although six studies reported regional alterations. While changes in the hippocampus and parahippocampus were frequently identified, the findings were inconsistent across studies
Loría F, Petrosino S, Hernangomez M, Mestre L, Spagnolo A, Correa F, Di Marzo V, Docagne F, Guaza C. 2010. “An endocannabinoid tone limits excitotoxicity in vitro and in a model of multiple sclerosis” Neurobiol Dis 37(1): 166-76
http://www.ncbi.nlm.nih.gov/pubmed/19815071/
we observed that neuronal cells respond to excitotoxic challenges by the production of endocannabinoid molecules which in turn exerted neuroprotective effects against excitotoxicity. The inhibitor of endocannabinoid uptake, UCM707, protected specifically against AMPA-induced excitotoxicity, by activating CB(1) and CB(2) cannabinoid receptors, as well as the nuclear factor, PPARgamma. This neuroprotective effect was reverted by blocking the glial glutamate transporter, GLT-1
enhancement of the endocannabinoid tone leads to neuroprotection against AMPA-induced excitotoxicity and provides therapeutic effects in this model of multiple sclerosis
Lu T, Newton C, Perkins I, Friedman H, Klein T. 2006. “Cannabinoid treatment suppresses the t-helper cell-polarizing function of mouse dendritic cells stimulated with Legionella pneumophila infection” Pharmacology 319(1): 269-276
http://jpet.aspetjournals.org/content/319/1/269.long
the immunization potential of the Lp-loaded cells along with Th1 cytokine production was attenuated by THC treatment at the time of in vitro infection. In addition, THC-treated and Lp-loaded DCs were poorly stimulated in culture-primed splenic CD4+ T cells to produce interferon-γ; however, this stimulating deficiency was reversed by adding recombinant interleukin (IL)-12p40 protein to the cultures. Moreover, THC treatment inhibited the expression of DC maturation markers, such as major histocompatibility complex class II and costimulatory molecules CD86 and CD40 as determined by flow cytometry and suppressed the Notch ligand, Del-ta4, as determined by reverse transcription-polymerase chain reaction. However, THC treatment did not affect other DC functions, such as intracellular killing of Lp, determined by colony-forming unit counts of bacteria, and Lp-induced apoptosis, determined by annexin V staining. In conclusion, the data suggest that THC inhibits Th1 activation by targeting essential DC functions, such as IL-12p40 secretion, maturation, and expression of costimulatory and polarizing molecules
Lukas S, Mendelson J, Benedikt R. 1995. “Electroencephalographic correlates of marihuana-induced euphoria” Drug Alcohol Depend 37(2): 131-40
http://www.ncbi.nlm.nih.gov/pubmed/7758402
Euphoria while plasma THC rapidly rising correlated with EEG alpha power
Lull M, Block M. 2010. “Microglial activation & chronic neurodegeneration” Neurotherapeutics 7(4): 354-365
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2951017/
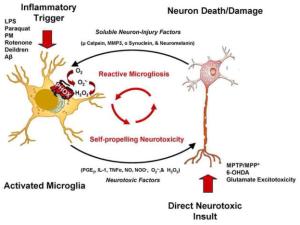
Neurodegenerative diseases (ex. Alzheimer’s Disease, Parkinson’s Disease, Huntington’s Disease, ALS, etc.) share many common characteristics, such as changes in microglial number and morphlogy, elevated cytokine levels, oxidative stress, and progressive neuronal loss. Increasing evidence reports that microglia can become a chronic source of cytokines and reactive oxygen (ROS) to drive progressive neuron damage and are implicated in the chronic nature of neurodegenerative diseases
Microglia reside in the CNS, comprise approximately 12% of the brain2
the original microglial population in the CNS differentiates from cells of the myeloid lineage that originate in the bone marrow2, which occurs early in embryonic development.3 These myeloid cells can be found within the CNS by embryonic day 8 in rodents4, and by the 12th week of gestation in humans.5
studies using bone marrow-chimeras have shown that circulating monocytes infiltrate the CNS under different conditions of injury, inflammation, or disease.9–11
one role of microglia is to act as the brain’s immune defense against disease and injury. In addition to these duties, however, microglia are involved in number of processes in the normal, healthy CNS.
In [resting] state, microglia take on a ramified appearance, where long thin processes extend from the cell body and into the surrounding milieu. The immunological phenotype of this state is characterized by low expression of major histocompatability complex (MHC) proteins and other antigen presenting surface receptors.18This phenotype is in stark contrast to that of other macrophages, which exist in a more readily activated state
Despite the fact that these ramified microglia are referred to as resting, they are constantly surveying the surrounding environment by extending and retracting their processes.23, 24 In doing this, microglia are able to sample the microenvironment, maintain homeostasis, and identify signals that require a response. When reacting to extracellular signals, such as the presence of pathogens, foreign material and dead or dying cells, microglia may undergo a morphological change into an ameboid shape with short or non-existent processes.25 … result in changes in surface receptor expression, the release of pro- or anti-inflammatory factors, recruitment molecules, and ROS, among others.26–29
Microglia are activated in response to brain injuries and immunological stimuli25, 30–32 to undergo dramatic alterations in morphology, changing from resting, ramified microglia into activated, amoeboid microglia
For microglia surrounding neurons, subtypes of microglia can provide trophic support to neurons through the release of nerve growth factors, neurotrophins, and other neurotrophic factors.34 These microglia are also capable of assisting in synaptic plasticity, an observation that was first made in the mid-20th century.35 Along these lines, microglia have been implicated as the “brains electricians”,36 where the release of neurotrophic factors and anti-inflammatory cytokines from microglia has been shown to promote synaptic plasticity.37–39 Specifically in response to injury, activated microglia have been shown to surround damaged neurons and participate in synaptic stripping, a process of removing branches from damaged neurons to promote repair and regrowth,35, 40, 41 although recent evidence shows that microglia may play an indirect, anti-inflammatory role in this process.42 Notably, these beneficial microglial functions often involve changes resembling an activated morphology and protein expression, yet the function is distinct from a classic pro-inflammatory response
In fact, the majority of microglial functions are beneficial and necessary for a healthy CNS, as activated microglia are critical for CNS wound healing.36… evidence supports that when microglia become neurotoxic, this is due to both the loss of the beneficial functions36 and/or a shift to a pro-inflammatory phenotype.1, 45, 46
Microglia are readily activated by an extensive list of pro-inflammatory stimuli, such as lipopolysaccharide (LPS),49–51 pesticides (ex. Paraquat,52 dieldrin,53, 54 lindane,54 and rotenone55), disease proteins (ex. Beta-amyloid (Aβ),56 α synuclein, 57 and HIV-Tat58), air pollution,59 and even neuron damage.46 In fact, it has been proposed that many disease proteins and environmental toxicants trigger a toxic microglial response because they are misinterpreted as a pathogen.1 In response to these triggers, microglia can produce of a large array of cytotoxic factors, such as superoxide (O2•−),27 nitric oxide (NO),60, 61 tumor necrosis factor alpha (TNFα)62, 63 and inflammatory prostaglandins.64
While the microglial superoxide response is immediate, microglia also respond to LPS with a later pulse of pro-inflammatory factors, such as TNFα, IL1-β, and NO, both in vivo and in vitro.65 Specifically, there is a delay in the production of TNFα, NO, prostaglandin E2 (PGE2) and IL-1β, where TNF-α peaks at 6 hours, NO peaks at 12 hours and both PGE2 and IL-1β peak at approximately 24 hours.65 Thus, one mechanism through which microglia are thought to cause neuron damage is through the excessive and inappropriate release of toxic factors
Neurodegenerative diseases are characterized by chronic and progressive neuronal loss, pathological levels of cytotoxic substances such as extracellular debris, elevated levels of pro-inflammatory factors, and production of reactive oxygen species, resulting in oxidative stress
not only is β-amyloid directly toxic to neurons,79 but it also causes microglia to cluster around plaques and become activated, which may perpetuate neuronal damage and death.80, 81
a critical role for neuroinflammation in dopaminergic (DA) neuron damage, as cytokines such as TNFα106–108 have been shown to exert DA neuron damage. In fact, continuous expression of low levels of TNFα in the SN induced by an adenoviral vector will cause chronic microglia/macrophage activation, progressive neurodegeneration, and delayed motor symptoms.109
Specific to microglia, ROS produced from NADPH oxidase has been shown to mediate changes in microglia morphology, 130 pro-inflammatory gene expression,130 and upregulation of markers in response to immunological stimuli.136
NADPH oxidase has also been linked to microglia-derived oxidative stress from a variety of neurotoxic insults, such as rotenone, 126DEP,127 α-synuclein,57 Aβ,128 PQ,139 dieldrin,54 DA neuronal injury,137, 140 prothrombin kringle-2,141 β2 adrenergic receptor activation,142 angiotensin,143 and cerebral ischemia-reperfusion injury,144, 145 indicating that microglial NADPH oxidase activation may also be a common denominator of microglial activation associated with neurotoxicity
these cells are also more likely to establish chronic pro-inflammatory responses, rather than demonstrate resolution of the innate immune response, as is common in the peripheral immune system.156
While microglial activation was initially perceived as a transient event, 31 it is now thought to be chronic and culpable in the propagation of disease.1, 94, 157 Reports have shown that microglia can remain chronically activated140, 158, 159 in a process that has been termed reactive microgliosis. Reactive microgliosis can be defined as microglial activation that occurs in response to neuronal damage, which is then perpetuated by further microglial activation and neurotoxicity (). Thus, a self-propelling and progressive cycle of microglial activation and neuron damage ensues.1
upon injury with the DA neurotoxin MPP+, DA neurons released soluble neuron injury factors that activated microglia and were selectively toxic to DA neurons in mixed mesencephalic neuron-glia cultures through NADPH oxidase.166
during critical periods of embryonic development (E 10.5), maternal exposure to low concentrations of LPS in mice impacts microglial activation and DA neuron survival in offspring that persists into adulthood.170, 171
not only can microglia induce neuron damage, but that microglia can become persistently activated to produce continuous and uncontrolled neurotoxicity that fails to resolve, long after the instigating stimulus has dissipated
The phenomenon of microglial priming offers valuable insight into why microglia continue to respond to additional stimuli in the chronic cycle of neuroinflammation (). In the case of priming, microglia are not just exhibiting and enhanced toxic microglial response. Rather, in the case of priming, the microglial phenotype shifts, where a much lower stimulus is needed to exact a toxic microglial response, enhancing the probability that the chronic cycle of toxic reactive microgliosis will continue
By altering concentrations of intracellular ROS and consequent redox-signaling, NADPH oxidase is reported to prime the microglial response to further insult. Triggers of microglia activation, such as rotenone55 and neuronal death,179 are shown to prime microglia through NADPH oxidase and result in synergistic microglial activation that is associated with neurotoxicity upon additional insult
Lüscher C, Slesinger P. 2010. “Emerging concepts for G protein-gated inwardly rectifying potassium (GIRK) channels in health and disease” Nat Rev Neurosci 11(5): 301-315
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3052907/
Much of the interplay of excitatory and inhibitory signals required for normal neuronal function occurs in the dendrites of neurons1. Detailed electron and light microscopic studies show that fast excitatory inputs are mediated by ionotropic glutamate receptors such as NMDARs and AMPARs that are located in the postsynaptic density at the head of the spine, whereas inhibitory synapses typically form on the soma and dendritic shafts1. The fast inhibitory signals are mediated by ionotropic GABAA/glycine receptors. In addition to this fast inhibition, a slower inhibitory postsynaptic potential (sIPSC) exists that is mediated by G protein-activated inwardly rectifying potassium (GIRK or Kir3) channels that are located perisynaptically outside the postsynaptic density in the spine and also on the shaft
GIRK channels reside in a macromolecular signaling complex, governed by protein-protein interactions, limited diffusion and lipid domains
chronic stimulation of endogenous M2 muscarinic (M2R) receptors leads to down regulation of both the receptors and GIRK channels
In hippocampus, GIRK channels are expressed on the dendritic shafts of hippocampal neurons… tight association of GIRK channels and GABAB receptors on dendritic spines
In the cerebellum, immunoelectron microscopy studies show that GIRK2/GIRK3 channels are localized in the postsynaptic density, while GIRK1/GIRK3 channels are perisynaptic112. It is tempting to speculate that the subcellular localization may correlate with the Gβγ affinity (GIRK2/GIRK3; lower EC50) in neurons.
Additional studies will need to clarify the role of collision coupling and whether changes in GIRK signaling occur at the level of the GPCR/GIRK macromolecular complex or individual components in the signaling pathway.
The physiological activation of GIRK channels can shape the neuronal network behavior in many areas of the brain at different levels. The basal activity of GIRK channels contributes to the resting potential of neurons, shifting the membrane voltage by ~−8 mV27. This hyperpolarization of the resting membrane potential decreases electrical excitability. In addition, receptor activation of GIRK channels provides another level of inhibition, to which three different changes in signaling can be generally ascribed.
a train of action potentials fired by low threshold spiking (LTS) interneurons of the cortex results in long-lasting hyperpolarization. This change in excitability occurs in a cell-autonomous way through endocannabinoids released from dendrites and the activation of CB1 receptors coupled to GIRK channels on the very same dendrites113. This modulation of LTS neurons may lead to long-lasting changes in cortical networks owing to altered glutamate transmission in pyramidal neurons. Dopamine neurons also exhibit a form of autaptic inhibition. Here, dendodendritic release of dopamine activates dopamine D2 receptors coupled to GIRK channels, leading to suppression of firing
The slow time course of outward current through GIRK channels therefore correlates with the time required for diffusion of the neurotransmitter and is not limited by the speed of G protein activation
In midbrain DA neurons, slow activation of GIRKs via D2 receptors originates from local pooling of dendrodendritically released DA
recent characterizations of GABAB receptor mediated presynaptic inhibition has revealed a component that is sensitive to a GIRK-specific channel inhibitor tertiapin123, suggesting a presynaptic role for GIRK channels in inhibiting neurotransmitter release2,124,125. Thus, GABA may inhibit pre-synaptic release concurrent with postsynaptic hyperpolarization
endogenous adenosine may suppress gamma oscillations in the hippocampus127, possibly due to the selective expression of A1 receptors on pyramidal neurons, which also activate GIRK channels
activation of NMDA receptor in cultured dissociated hippocampal neurons increases the level of the surface expression of GIRK1 and GIRK2 channel in the soma and dendrites within minutes
Increased expression of GIRK channels would reduce excitability, dampening signaling of hippocampal pyramidal neurons. For example, GIRK channel activation is implicated in the maintenance of LTP at glutamatergic synapses
Less is known concerning the mechanism of this plasticity but could involve desensitization of D2 receptors, modulation of G-protein function and/or GIRK channel availability (as GIRK channels can rapidly redistribute)
direct application of opioids into the PAG inhibits GABA neurons through both GIRK-independent (presynaptic) and GIRK-dependent (postsynaptic) mechanisms, which is sufficient to induce analgesia
suggests that GIRK channels ‘gate’ the induction of opioid dependence by silencing the neurons in the locus coeruleus every time opioids are injected
loss of GIRK function can lead to excessive excitability leading, such as in epilepsy, while a gain of GIRK function can significantly reduce neural activity, as postulated in Down’s syndrome. Second, loss of selectivity can cause ion fluxes across GIRK channels, such as excessive K+ efflux that triggers cell death, exemplified in a model of Parkinson’s disease
GIRK2 knockout mice develop spontaneous convulsions and show a propensity for generalized seizures when injected with a pro-convulsive GABAA receptor antagonist
electroconvulsive shock leads to increased expression of GIRK channels
chromosome 21 (duplicated in Down’s syndrome) contains the Girk2 (Kcnj6) gene
genetic upregulation of girk2: hippocampal LTP is reduced while LTD is enhanced in both lines and therefore synaptic plasticity of glutamatergic transmission is altered
Opioids and the club drug γ-hydroxbutyrate (GHB) activate GIRK channels leading to disinhibition of DA neurons. Morphine stimulates μ-opioid receptors that are selectively expressed on interneurons of the VTA and activate GIRK channels, reducing the firing of these cells, eventually leading to disinhibition of DA neurons153. The effect of GHB is more complex, as GABAB receptors are expressed on both GABA and DA neurons. However, DA neurons in the VTA are an order magnitude less sensitive (higher EC50) to GABAB receptor agonists than GABA interneurons43most likely due to the selective expression of GIRK2c and GIRK3 and the lack of GIRK1 subunit (see discussion above). Consequently, low concentrations of GHB only activate GIRK currents in GABA neurons (containing GIRK1 subunits), leading to disinhibition of DA neurons
In the case of cannabinoids, the disinhibition of DA neurons seems to occur primarily through presynaptic, GIRK-independent mechanisms
he weaver mouse132, where constitutively active GIRK2wv channels produce chronic depolarization and cell death in a subset of neurons in the brain
important to note that the suggested roles for GIRK channels in brain diseases are based on observations in animal models. Future genetic studies may provide direct evidence for altered GIRK function in human diseases
Lumley M, Harkness K. 2007. “Specificity in the Relations among Childhood Adversity, Early Maladaptive Schemas, and Symptom Profiles in Adolescent Depression” Cognitive Therapy and Research 31(5): 639-657
http://link.springer.com/article/10.1007%2Fs10608-006-9100-3
schemas with themes of loss/worthlessness preferentially mediated the relation between childhood adversity and anhedonic symptoms, while schemas with themes of danger preferentially mediated the relation between childhood adversity and anxious symptoms. Sexual abuse was not significantly associated with either depression or anxiety symptoms
Lundberg D, Daniel A, Thayer S. 2005. “Delta(9)-Tetrahydrocannabinol-induced desensitization of cannabinoid-mediated inhibition of synaptic transmission between hippocampal neurons in culture” Neuropharmacology 49(8): 1170-7
http://www.ncbi.nlm.nih.gov/pubmed/16157354
synthetic full CB1 agonist Win55,212-2 produces much greater CB1 desensitization than does THC
Lutz B, Marsicano G, Maldonado R, Hillard C. 2015. “The endocannabinoid system in guarding against fear, anxiety and stress” Nature Reviews Neuroscience 16: 705-718
http://www.nature.com/nrn/journal/v16/n12/full/nrn4036.html
Lyketsos C, Garrett E, Liang K-Y, Anthony J. 1999. “Cannabis uses and cognitive decline in persons under 65 years of age” Am J Epidemiol 149(9): 794-800
http://aje.oxfordjournals.org/content/149/9/794
The purpose of this study was to investigate possible adverse effects of cannabis use on cognitive decline after 12 years in persons under age 65 years… 1,318 participants in the Baltimore, Maryland, portion of the Epidemiologic Catchment Area study who completed the Mini-Mental State Examination (MMSE) during three study waves in 1981, 1982, and 1993–1996
After 12 years, study participants’ scores declined a mean of 1.20 points on the MMSE (standard deviation 1.90), with 66% having scores that declined by at least one point. Significant numbers of scores declined by three points or more (15% of participants in the 18–29 age group). There were no significant differences in cognitive decline between heavy users, light users, and nonusers of cannabis. There were also no male-female differences in cognitive decline in relation to cannabis use. The authors conclude that over long time periods, in persons under age 65 years, cognitive decline occurs in all age groups. This decline is closely associated with aging and educational level but does not appear to be associated with cannabis use
Lynch M, Ware M. 2015. “Cannabinoids for the treatment of chronic non-cancer pain: an updated systematic review of randomized controlled trials” J Neuroimmune Pharmacol 10(2): 293-301
http://www.ncbi.nlm.nih.gov/pubmed/25796592
Eleven trials published since our last review met inclusion criteria. The quality of the trials was excellent. Seven of the trials demonstrated a significant analgesic effect. Several trials also demonstrated improvement in secondary outcomes (e.g., sleep, muscle stiffness and spasticity). Adverse effects most frequently reported such as fatigue and dizziness were mild to moderate in severity and generally well tolerated. This review adds further support that currently available cannabinoids are safe, modestly effective analgesics that provide a reasonable therapeutic option in the management of chronic non-cancer pain
Lynskey M, Glowinski A, Todorov A, Bucholz K, Madden P, Nelson E, Statham D, Martin N, Heath A. 2004. “Major depressive disorder, suicidal ideation, and suicide attempt in twins discordant for cannabis dependence and early-onset cannabis use” Arch Gen Psychiatry 61(10): 1026-32
https://www.ncbi.nlm.nih.gov/pubmed/15466676/
Individuals who were cannabis dependent had odds of suicidal ideation and suicide attempt that were 2.5 to 2.9 times higher than those of their non-cannabis-dependent co-twin. Additionally, cannabis dependence was associated with elevated risks of MDD in dizygotic but not in monozygotic twins. Those who initiated cannabis use before age 17 years had elevated rates of subsequent suicide attempt (odds ratio, 3.5 [95% confidence interval, 1.4-8.6]) but not of MDD or suicidal ideation. Early MDD and suicidal ideation were significantly associated with subsequent risks of cannabis dependence in discordant dizygotic pairs but not in discordant monozygotic pairs.
Comorbidity between cannabis dependence and MDD likely arises through shared genetic and environmental vulnerabilities predisposing to both outcomes. In contrast, associations between cannabis dependence and suicidal behaviors cannot be entirely explained by common predisposing genetic and/or shared environmental predispositions. Previously reported associations between early-onset cannabis use and subsequent MDD likely reflect shared genetic and environmental vulnerabilities, although it remains possible that early-onset cannabis use may predispose to suicide attempt
While our results are consistent with the hypothesis that early cannabis use increases risks for subsequent suicide attempt, we cannot exclude the possibility that both these outcomes result from aspects of the nonshared environment that were not controlled in our analyses. In addition to the pharmacological effects of cannabis, there are a number of other potential mechanisms that may underlie any association between cannabis use and risks of suicidal behaviors. In particular, the social context within which cannabis is used and obtained may also promote access to other drugs27 and a range of life events and circumstances, including risks of legal sanctions against possession and use of cannabis,30 that may predispose cannabis users to suicidal behaviors
Lyons M, Bar J, Panizzon M, Toomey R, Eisen S, Xian H, Tsuang M. 2004. “Neuropsychological consequences of regular marijuana use: a twin study” Psychol Med 34(7):1239-50
http://www.ncbi.nlm.nih.gov/pubmed/15697050
50 domains tested, only one difference found “only the performance of the block design subtest of the Wechsler Adult Intelligence Scale–Revised reached a level of statistical significance.”
“The results indicate an absence of marked long-term residual effects of marijuana use on cognitive abilities.”
MacMillan H, Boyle M, Wong M, Duku E, Fleming J, Walsh C. 1999. “Slapping and spanking in childhood and its association with lifetime prevalence of psychiatric disorders in a general population sample” CMAJ 161(7): 805-809
http://www.cmaj.ca/content/161/7/805.full
n= 9953, “sometimes” slapped or spanked (33.4%) or “rarely” (40.9%); 5.5% reported that this occurred “often.” The remainder (20.2%) reported “never”
“often” or “sometimes” had significantly higher lifetime rates of anxiety disorders (adjusted odds ratio [OR] 1.43, 95% confidence interval [CI] 1.04-1.96), alcohol abuse or dependence (adjusted OR 2.02, 95% CI 1.27-3.21) and one or more externalizing problems (adjusted OR 2.08, 95% CI 1.36-3.16)
Macleod J, Oakes R, Copello A, Crome I, Egger M, Hickman M, Oppenkowski T, Stokes-Lampard H, Smith G. 2004. “Psychological and social sequelae of cannabis and other illicit drug use by young people: a systematic review of longitudinal general population studies” The Lancet 363(9421): 1579-1588
http://www.thelancet.com/journals/lancet/article/PIIS0140-6736(04)16200-4/abstract
Use of illicit drugs, particularly cannabis, by young people is widespread and is associated with several types of psychological and social harm. These relations might not be causal. Causal relations would suggest that recreational drug use is a substantial public health problem. Non-causal relations would suggest that harm-reduction policy based on prevention of drug use is unlikely to produce improvements in public health.
48 relevant studies, of which 16 were of higher quality and provided the most robust evidence. Fairly consistent associations were noted between cannabis use and both lower educational attainment and increased reported use of other illicit drugs. Less consistent associations were noted between cannabis use and both psychological health problems and problematic behaviour. All these associations seemed to be explicable in terms of non-causal mechanisms
Available evidence does not strongly support an important causal relation between cannabis use by young people and psychosocial harm, but cannot exclude the possibility that such a relation exists. The lack of evidence of robust causal relations prevents the attribution of public health detriments to illicit drug use
Madrigal J, Garcia-Bueno B, Caso J, Perez-Nievas B, Leza J. 2006. “Stress-induced oxidative changes in brain” CNS Neurol Disord Drug Targets 5(5): 561-8
http://www.ncbi.nlm.nih.gov/pubmed/17073658/
Some of the most impressive effects of the stress in brain are the atrophy of hippocampal dendrites or even the reduction of the hippocampal size observed in brains from subjects exposed to severe or chronic stress. Obviously, before reaching this point of damage there are many other processes taking place in the stressed CNS. The release of glucocorticoids is one of the first features of the stress response. Glucocorticoids can result in neurotoxicity through different mechanisms, including modifications in the energy metabolism or via an increase in excitatory amino acids such as glutamate in the extracellular space. Glutamate can induce neuronal excitotoxicity. This sequence of events leads to the activation of TNFalpha convertase (TACE) and TNFalpha release in brain of rats subjected to restraint stress. One of the multiple effects exerted by this cytokine is to initiate the translocation of the transcription factor NFkappaB to neuronal nuclei. NFkappaB activation results in the induction of iNOS and COX2, two enzymes responsible for a great portion of the neurological damage produced in models of stress
Maes M, Song C, Lin A, De Jongh R, Van Gastel A, Kenis G, Bosmans E, De Meester I, Benoy I, Neels H, Demedts P, Janca A, Scharpe S, Smith R. 1998. “The effects of psychological stress on humans: increased production of pro-inflammatory cytokines and a Th1-like response in stress-induced anxiety” Cytokine 10(4): 313-8
http://www.ncbi.nlm.nih.gov/pubmed/9617578
Psychological stress significantly increased the stimulated production of tumour necrosis factor alpha (TNF-alpha), interleukin 6 (IL-6), IL-1 receptor antagonist (IL-1Ra), interferon gamma (IFN-gamma) and IL-10. Students with high stress perception during the stressful condition had a significantly higher production of TNF-alpha, IL-6, IL-1Ra and IFN-gamma than students with a low-stress perception. Students with a high anxiety response had a significantly higher production of IFN-gamma and a lower production of the negative immunoregulatory cytokines, IL-10 and IL-4, than students without anxiety. These findings suggest that, in humans, changes in the production of the pro-inflammatory cytokines, TNF-alpha, IL-6 and IFN-gamma, and negative immunoregulatory cytokines, IL-10 and IL-4, take part in the homeostatic responses to psychological stress and that stress-induced anxiety is related to a T-helper-1-like response
Maes M, Berk M, Goehler L, Song C, Anderson G, Galecki P, Leonard B. 2012. “Depression and sickness behavior are Janus-faced responses to shared inflammatory pathways” BMC Medicine 10: 66
http://www.biomedcentral.com/1741-7015/10/66
It is of considerable translational importance whether depression is a form or a consequence of sickness behavior. Sickness behavior is a behavioral complex induced by infections and immune trauma and mediated by pro-inflammatory cytokines. It is an adaptive response that enhances recovery by conserving energy to combat acute inflammation. There are considerable phenomenological similarities between sickness behavior and depression, for example, behavioral inhibition, anorexia and weight loss, and melancholic (anhedonia), physio-somatic (fatigue, hyperalgesia, malaise), anxiety and neurocognitive symptoms. In clinical depression, however, a transition occurs to sensitization of immuno-inflammatory pathways, progressive damage by oxidative and nitrosative stress to lipids, proteins, and DNA, and autoimmune responses directed against self-epitopes. The latter mechanisms are the substrate of a neuroprogressive process, whereby multiple depressive episodes cause neural tissue damage and consequent functional and cognitive sequelae. Thus, shared immuno-inflammatory pathways underpin the physiology of sickness behavior and the pathophysiology of clinical depression explaining their partially overlapping phenomenology. Inflammation may provoke a Janus-faced response with a good, acute side, generating protective inflammation through sickness behavior and a bad, chronic side, for example, clinical depression, a lifelong disorder with positive feedback loops between (neuro)inflammation and (neuro)degenerative processes following less well defined triggers
The characteristic behavioral pattern consists of malaise, hyperalgesia, pyrexia, listlessness and disinterest in social interactions with the environment, lethargy, behavioral inhibition, reduction of locomotor activity, exploration and grooming, reduction of reproductive performance, anhedonia, somnolence and sleepiness, anorexia and weight loss, failure to concentrate, and anxiety. There is evidence that sickness behavior is mediated through the effects of pro-inflammatory cytokines (PICs), such as IL-1, TNFα and IL-6 [3–10].
clinical depression is an immuno-inflammatory disorder characterized by among other things increased levels of PICs and acute phase proteins, including C-reactive protein and haptoglobin [11–20]
‘vegetative symptoms’ of depression, such as anorexia, weight loss, and psychomotor retardation, are significantly associated with inflammatory markers in clinical depression, such as increased levels of plasma haptoglobin, an acute phase protein, synthesis of which is induced by the three abovementioned PICs [1, 2]
O’Mahony S, Clarke G, Borre Y, Dinan T, Cryan J. 2015. “Serotonin, tryptophan metabolism and the brain-gut-microbiome axis” Behavioural Brain Research 277 32-48
http://www.ncbi.nlm.nih.gov/pubmed/25078296 in dropbox
The brain-gut axis is a bidirectional communication system between the central nervous system and the gastrointestinal tract. Serotonin functions as a key neurotransmitter at both terminals of this network
substantial overlap between behaviours influenced by the gut microbiota and those which rely on intact serotoner-gic neurotransmission
developing serotonergic system may be vulnerable to differential microbial colonisation patterns prior to the emergence of a stable adult-like gut microbiota
ability of the gut microbiota to control host tryptophan metabolism along the kynurenine pathway, thereby simultaneously reducing the fraction available for serotonin synthesis and increasing the production of neuroactive metabolites. The enzymes of this pathway are immune and stress-responsive
neural processes in the gastrointestinal tract which can be influenced by local alterations in serotonin concentrations with subsequent relay of signals along the scaffolding of the brain-gut axis to influence CNS neurotransmission
the vast major-ity of serotonin is located in the gut where it is synthesised from tryptophan in the enterochromaffin cells (ECs) of the gastrointesti-nal tract and is also present in enteric nerves
the dominant physiological pathway for tryptophan is actually along the kynurenine pathway (see Fig. 2) [2,17] . Kynurenine is produced from tryptophan by the action of the largely hepatic based enzyme, tryptophan-2,3-dioxygenase (TDO) or the ubiquitous indoleamine-2,3-dioxygenase (IDO) [18] . TDO can be induced by glucocorticoids or indeed tryp-tophan itself whereas IDO is influenced by certain inflammatory stimuli, IFN-gamma being the most potent inducer [11]
Once kynurenine is produced, it is further metabolised along two distinct arms of the pathway with one leading to the production of the neuroprotective kynurenic acid (7 nicotinic acetylcholine receptor antagonist and N-methyl-d-aspartate (NMDA) receptor antagonist at glycine site) and the other to the neurotoxic quinolinic acid (a NMDA receptor agonist). Moreover, kynurenic acid, which can be neuro-protective against quinolinic acid induced excitotoxicity, can also induce cognitive impairment when abnormally elevated
circulating tryptophan concentrations do not solely determine CNS availability as it competes with other large neutral amino acids for transport across the BBB [25] and with exercise, for example, also influencing CNS distribution and utilisation
in vulnerable populations, reducing the circulating concentra-tions of tryptophan can impact functionally on mood, reinstating, as ATD does, depressive symptoms in patients who have success-fully responded to selective serotonin reuptake inhibitors (SSRIs)
The gut microbiome refers to the collection of microorganisms and their genomes in the gut habitat which are now regarded as a critical node in the brain-gut axis [5,9]. One outcome of the recent intense focus on this ‘virtual organ’ using metagenomic approaches is the realisation that microorganisms in our gastroin-testinal tract outnumbers the human cells in our bodies by a factor of 10 and contains 150 times as many genes as our genome
during the period follow-ing birth until the attainment of a stable adult-like microbiota at approximately 3 years of age, there is considerable flux which may prime for healthy in later life
infants born by caesarean section (C-section) have a distinct microbiota more akin to the skin microbiota of their mothers, including Staphy-lococcus , Corynebacterium and Propionibacterium spp [34] with a low representation of Bifidobacteria spp… preterm infants have a different microbiota (dominated by Proteobacteria initially and a lack of detectable Bifidobacterium and Lactobacillus genera)
abundance of certain Bifidobacterium spp in exclusively breast-fed… gut microbiota of formula-fed infants typically displays a lower prevalence of Bifidobacteria and a higher abundance of coliforms, Bacteroides and Clostridium dif-ficile
increased risk for psychopathology arises from detrimental colonisation patterns
although there is a high inter-individual gut microbiota variability, an adult individ-ual’s intestinal microbial community is relatively stable over time in the absence of any major ecological trauma… also exhibits plasticity dur-ing adulthood and is heavily influenced by nutritional factors and indeed potentially subject to rapid compositional shifts following
alterations in diet
gut microbiota (see Section 3) is essential for normal brain development and behaviour and can impinge on tryptophan and serotonin… germ-free condition is characterised by increased
plasma tryptophan concentrations which can be normalised following colonisation
Increased IDO activity has been demonstrated in both male and female IBS populations which may be related to low-grade immune changes in this disorder
certain bacterial strains harbour a tryptophanase enzyme that produces indole from tryptophan
The balance between bacterial tryptophan utilisation and metabolism, tryptophan synthesis, serotonin production and indeed the bacterial response to exogenous elevations in serotonin likely plays an important role in determining local gastrointestinal and circulating tryptophan availability for the host in addition to the dietary supply of this essential amino acid
Serotonin-producing bacterial strains: Lactococcus lactis subsp. cremoris; L. lactis subsp. lactis; Lactobacillus plantarum; Streptococcus thermophilus; Escherichia coli; Morganella morganii; Klebsiella pneumoniae; Hafnia alvei
Before the formation of synaptic interneuronal connections and the development of the BBB, neurons function as secretory cells so the developing brain has been considered as an early endocrine organ
5-HT is considered as an integral signalling molecule that regulates the development of many targets throughout the body
Although the BBB is func-tional at birth, the fragility of the developing cerebral vasculature indicates that the neonatal brain is more vulnerable to circulating toxins than during adulthood
The central serotonergic system originates in the raphe nuclei in the brainstem which project to virtually every part of the central nervous system and is implicated in modulating physiological processes such as mood, sleep, aggression and sexual behaviour
from very early stages 5-HT raphe cells appear to be subdivided into specific subsets, projecting to different brain areas and may require specific transcriptional and environmental factors for their development
In the human brain the levels of 5-HT remain fairly stable during normal aging yet overall 5-HT receptors appear to reduce by about 30–50% over the lifespan
serotonin released from the EC cells mediates many gastroin-testinal functions, including secretion, peristalsis, vasodilation, perception of pain and nausea through the activation of the 5-HT receptors (see Table 2) which are found on intrinsic and extrinsic afferent nerve fibres
Despite the ENS being referred to as the “little brain” or the “sec-ond brain” because of its autonomy, it is still well connected to the CNS via motor and sensory fibres of the sympathetic and parasym-pathetic nervous systems
early-developing enteric neurons or their neurotransmitters might influence the fate of later ones due to the sequential developmental order
stimulation of 5-HT4 receptors protects enteric neurons against apoptotic death and inhibits inflammation induced axon terminal degeneration and autophagy [118] . 5-HT4 receptors are also involved in the mobilisation of extraganglionic adult stem cells to form new neurons that may replace damaged or killed ones [123] . The development of microvilli, which increase the surface area for efficient absorption of nutrients, is induced by 5-HTP, the 5-HT precursor, but not 5-HT
The immune system forms an integral component of the signalling pathway in the brain-gut axis
Gut associated lymphoid tissue, which modulates tolerance versus luminal antigens and prevents the tran-sit of potentially harmful antigens across the intestinal barrier, is dependent on the microbiota for development [129] . The immature immune system of the newborn has a Th2 bias which shifts to a Th1 response with bacterial colonisation [125] . Different lactic acid producing bacteria such as Bifidobacterium and Lactobaccillus have been shown to promote a Th1 shift
early-life antibiotic exposure is associated with allergic disease [132,133] and inflammatory bowel disease
Serotonin can modulate the immune response and hence poten-tially influence intestinal inflammation [135] . Several of the 5-HT receptors have been associated with immune cells such as lympho-cytes, monocytes, macrophages and dendritic cells which indicates that 5-HT plays an immune-modulatory role
Changes to the gastrointestinal content of 5-HT and EC cell population are associated with Crohns disease and ulcerative coli-tis [139–141] . Altered 5-HT signalling in both the central nervous system and the gastrointestinal tract have been implicated in IBS which may give rise to some of the intestinal and extra-intestinal symptoms seen in this disorder [142] . This is also suggested by the therapeutic effects of tricyclic antidepressants and SSRIs [143] as well as 5-HT4 receptor agonists and 5-HT3 receptor antagonists in the treatment of IBS symptoms
Vagal innervation is the major two way neural connection between the brain and the gut providing both motor and sensory innervation for numerous functions (e.g. satiety, nausea, sensation of visceral pain, sphincter operation and peristalsis)
The main stress axis, the hypothalamic pituitary adrenal (HPA) axis, provides hormonal communication within the brain-gut axis.
If this stress hypo-responsive period (thought to extend throughout childhood in humans) is maintained, it is evident that the stress system develops normally and the individual responds suitably to stressors as an adolescent and adult. In con-trast, if this period is interrupted through low-quality of parental care or abuse the axis can develop to over-activate in stressful situations
It is important to note that stress during this period can also modulate the sympathetic nervous system with associated functional interactions between the adrenal medulla and cortex [153–155] and potential implications for the CNS serotonergic sys-tem [156] . Indeed, bidirectional sympathetic pathways are also an important communication route of the brain-gut axis
Early-life stressors are associated with increased anxiety and depressive-like behaviours as well as GI disorders that have a stress component [52,159]. Given the influence of stress on the gut-brain axis [10] , appropriate development of the HPA axis is essential to the balanced functioning gut-brain axis. Stress has long been known to influence the composition of the gut microbiota [160] and we have shown that stress in early life alters the gastrointesti-nal bacterial content of the adult rat [161] . Hence, early life stress capable of activating the HPA axis can impact on the developing microbiota and vice versa, ultimately leading to an imbalance in the gut microbiota and an inappropriate stress response [97]
stress during this vulnerable period leads to dysfunction within several of the intertwined components of the gut-brain axis and increases susceptability to disease
Germ-free mice exposed to restraint stress exhibited a significantly higher level of adrenocorticotropic hormone (ACTH) and corticosterone than con-ventionally colonised mice [163]
Activation of the HPA axis during early life has been associated with an altered serotonergic system [164] . Maternal separation resulted in changes to 5-HT metabolism and attenuated expression of 5-HT1A and 5-HT2A receptors in various brain regions… stressed rats showed an increase in activated 5-HT positive neurons on the dorsal raphe nucleus, an area associated with the descending modulation of pain
appropriate development of both the brain-gut-microbiome axis and the 5-HT system depend on a balanced and stress-free environment
Germ-free mice robustly and reproducibly display less anxiety-like behaviours than their conventionally colonised counterparts… germ-free rats display increased anxiety-like behaviours
the developmental origins of anx-iety is closely associated with the developing serotonergic system [171]
in healthy volunteers, the latter combination of pro-biotic strains alleviated psychological distress including an index of depression
The mechanism of antidepressant action of minocycline could be related to neuroprotection, suppression of microglial activa-tion or anti-inflammatory actions
Further evidence of cognitive alterations in IBS relates to hippocampal mediated vis-uospatial memory deficits which appear to be related to indices of HPA axis function [193] . IBS patients also show impairments on a test of cognitive flexibility and have abnormal brain activ-ity in frontal brain regions during this task
intake of a fermented milk product with a probiotic can alter brain activity in regions of relevance to cognitive performance and which are innervated by serotonergic projections [196
Altered signalling from the gut is also proposed as being responsible for visceral hypersensitivity with immune activation, for example, causing sensitisation of neuronal afferents [203] and neuroplastic changes in the ENS
involve-ment of the microbiota in pain signalling extends beyond the gut with a full and balanced microbial community necessary for the appropriate development of pain signalling from the body [106] . Moreover, consumption of L. casei Shirota leads to a significant improvement in gastrointestinal symptoms, including abdominal pain, in Parkinson’s disease
factors interfering with the normal microbial ecology during this period may have deleterious effects in adulthood including those affecting appropriate development of pain pathways
high prevalence of anxiety and psycholog-ical disorders, have been shown to increase intestinal response to psychological stress [173]
bacteria also possess anxiolytic and antidepressant-like activity that can also affect this axis as well as the seroto-nergic system [61]
Candida, Streptococcus, Escherichia and Enterococcus are producers of 5-HT (Table 1) [84]
The correct functioning of the brain-gut-microbiome axis, which confers quality of life on an individual, depends on appropriate 5-HT signalling through the entire life span to nurture develop-ment, function and maintenance of this axis
Maier-Hein K, Brunner R, Lutz K, Henze R, Parzer P, Feigl N, Kramer J, Meinzer H-P, Resch F, Stieltjes B. 2014. “Disorder-specific white matter alterations in adolescent borderline personality disorder” 75: 81-88
http://www.ncbi.nlm.nih.gov/pubmed/23768862 in dropbox
Makela P, Wakeley J, Gijsman H, Robson P, Bhagwagar Z, Rogers R. 2005. “Low Doses of Delta-9 Tetrahydrocannabinol (THC) Have Divergent Effects on Short-Term Spatial Memory in Young, Healthy Adults” Neuropsychopharmacology 31: 462-470
We examined the effects of a single sublingual 5 mg dose of THC on a test of spatial working memory (requiring active manipulation of remembered spatial information for the management of future behavior) and a test of spatial span (requiring only the reproduction of sequences of previously presented spatial cues). In all, 19 healthy adults were administered 5 mg THC and placebo in a double-blind, placebo-controlled, within-subject, crossover design. Male participants performed more accurately than female participants. THC significantly enhanced spatial working memory performance of female participants. By contrast, male and female participants produced more intrusion errors during performance of the Spatial Span task. These results suggest that THC has relatively complex effects on spatial memory in human subjects
the contribution of DA to spatial working memory is mediated by the D1 (Arnsten et al, 1994; Sawaguchi and Goldman-Rakic, 1991) and D2 families of receptors (Arnsten et al, 1995; Zhang et al, 2004) that may influence mnemonic and response aspects of information processing within primate PFC (Wang et al, 2004; Williams and Goldman-Rakic et al, 1995). Other research attests to the importance of NA transmission, and the balanced activity of  1– and 2-adrenoceptors, in protecting the cognitive functions of the lateral PFC from the deleterious effects of stress (Arnsten, 2000; Arnsten and Contant, 1992; Arnsten and Jentsch, 1997; Birnbaum et al, 2000)
1– and 2-adrenoceptors, in protecting the cognitive functions of the lateral PFC from the deleterious effects of stress (Arnsten, 2000; Arnsten and Contant, 1992; Arnsten and Jentsch, 1997; Birnbaum et al, 2000)
2 healthy males and seven healthy females participated… One male and five female participants reported less than 10 lifetime uses of cannabis; eight males and two females reported between 10 and 100; and three males reported more than 100… mean age was 21.80.93 years (SE) and mean verbal IQ as estimated by the National Adult Reading Test (NART; Nelson, 1982) was 113.41.10.
THC extracted from standardized plants was administered in an ethanol and propylene glycol solution at 8.5 mg/ml concentration, using a pump-action sublingual spray… The total dose delivered was 5 mg THC in six actuations at 6-min intervals
HC significantly increased self-reported marijuana effects compared to placebo (1.300.46 vs 0.420.28) (F(1, 15)=9.32, p<0.01), with a trend for this effect to be maximal at +2 h onward (see Figure 1; M scale) (F(3, 45)=2.57, p=0.07). Sedative effects were also increased but not significantly so (Figure 1; PCAG scale) (F(1, 15)=2.90). There was no indication that THC induced euphoria in comparison with placebo (Figure 1; MBG scale) (F<1.00) or that it produced stimulant effects (Figure 1; BG and A scales) (both F’s<1.00). However, THC tended to enhance dysphoria and somatic effects (see Figure 1; LSD effects) (F(1, 15)=4.53, p=0.05), and particularly so in the female (2.450.54 to 3.550.37) compared to male participants (3.330.40 to 3.330.27) (F(1, 15)=4.53, p=0.05).
Treatment with 5 mg THC significantly reduced the number of between-search errors (ie number of returns to open a box in which a counter had already been found on the same trial) committed while performing the Spatial Working Memory task in comparison to placebo (3.090.70 vs 4.910.95) (F(1, 15)=8.92, p<0.01). This improvement was most apparent at the highest level of difficulty (F(2, 30)=5.82, p<0.01). In general, the effects of THC on the between-search errors were evident in female (7.571.53 vs 3.351.13) but not male participants (2.251.12 vs 2.830.82), so that the two-way interaction between treatment and gender was highly significant (F(1, 15)=15.57, p<0.005)
Within-search errors (ie the number of returns to boxes previously checked and shown to be empty earlier in the same search sequence) were not significantly altered after treatment with THC compared to treatment with placebo (0.240.09vs 0.110.51) (F(1, 15)=2.21)
no significant difference between overall strategy scores after THC compared to placebo treatment (26.931.51 vs 28.061.44) (F<1.00). However, female participants who completed the task first after placebo treatment showed improved strategy scores when completing the task again after THC (35.003.93 vs 26.504.12) compared to females who completed the task with the reverse treatment order (24.602.49 vs27.202.61)
The maximum memory span reached on the Spatial Span task was not significantly altered following THC treatment compared to placebo treatment (6.730.39 vs 7.010.27) (F<1.00)
the total number of errors was significantly reduced after THC treatment compared to placebo treatment in the male participants (F(1, 10)=6.92,p<0.05) but increased in the female participants (F(1, 5)=6.18, p=0.06)
pharmacological effects depend upon the level of baseline performance of participants under placebo, and are greatest in those whose baseline scores allow greater room for change (Kimberg and D’Esposito, 2003; Luciana et al, 1992; Milleret al, 1977).
The current results suggest that THC facilitates the actions of the dorsolateral PFC but impairs the actions of ventrolateral PFC
while THC did not reduce the maximum ‘span’ of correctly recalled sequences, it did compromise the representation of which cues belong in the sequence and which do not, thereby increasing intrusions when recalling sequences of even abstract, austere spatial cues of the kind used here. In summary, the present data suggest that low doses of THC have both beneficial and detrimental effects on the spatial memory of healthy human subjects, perhaps facilitating the manipulation of information but impairing the interstimulus associations that would otherwise promote accurate recall
Malfair A, Gallily R, Sumariwalla P, Malik A, Andreakos E, Mechoulam R, Feldmann M. 2000. “The nonpsychoactive cannabis constituent cannabidiol is an oral anti-arthritic therapeutic in murine collagen-induced arthritis” Proc Natl Acad Sci U S A 97(17): 9561-9566
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC16904/
CBD was administered after onset of clinical symptoms, and in both models of arthritis the treatment effectively blocked progression of arthritis. CBD was equally effective when administered i.p. or orally. The dose dependency showed a bell-shaped curve, with an optimal effect at 5 mg/kg per day i.p. or 25 mg/kg per day orally. Clinical improvement was associated with protection of the joints against severe damage
CBD, through its combined immunosuppressive and anti-inflammatory actions, has a potent anti-arthritic effect in CIA
Manrique-Garcia E, Zammit S, Dalman C, Hemmingsson T, Allebeck P. 2012. “Cannabis use and depression: a longitudinal study of a national cohort of Swedish conscripts” BMC Psychiatry 12: 112
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3480923/
A cohort study of 45 087 Swedish men
Only subjects with the highest level of cannabis use had an increased crude hazard ratio for depression (HR 1.5, 95% confidence interval (CI), 1.0-2.2), but the association disappeared after adjustment for confounders. There was a strong graded association between cannabis use and schizoaffective disorder, even after control for confounders, although the numbers were small (HR 7.4, 95% CI, 1.0-54.3).
De Marchi N, Petrocellis L, Orlando P, Daniele F, Fezza F, Marzo V. 2003. “Endocannabinoid signalling in the blood of patients with schizophrenia” Lipids Health Dis 2: 5
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC194767/
The amounts of anandamide were significantly higher in the blood of patients with acute schizophrenia than in healthy volunteers (7.79 ± 0.50 vs. 2.58 ± 0.28 pmol/ml). Clinical remission was accompanied by a significant decrease of the levels of anandamide (3.88 ± 0.72 pmol/ml) and of the mRNA transcripts for CB2 receptors and FAAH
changes in the blood levels of these endogenous mediators could explain in part, or be a consequence of, the modified immune response observed in the course of schizophrenia
the elevation of anandamide levels in our sample is confined to the acute phases of disease
no direct correlation between anandamide blood levels and the BPRS or CGI scores of each patient, be it before or after remission, was found. Our results suggest that an acute psychotic episode might be characterised, among other factors, by an elevation in the peripheral concentration of anandamide, thus possibly inducing a compensatory increased expression of the degrading enzyme, FAAH, in an attempt to normalise their circulating levels. It is of interest that our patients consistently showed the same pattern, with their level of circulating anandamide forming two well-identifiable and distinct clusters at around 8 and 3 pmol/ml for group 1 and 2, respectively
negative effect of leptin on endocannabinoid biosynthesis [26], and of the reduced levels of this hormone during acute schizophrenia [27], it is possible that reduced leptin causes the increased levels of blood anandamide observed in the schizophrenic patients investigated in the present study
it is possible that the reduced levels of anandamide following olanzapine treatment are a mere consequence of the reduced activity of lymphocytes and monocytes, which are normally capable of producing anandamide only after activation [19,20,29]
Márquez C, Poirier G, Cordero M, Larsen M, Groner A, Marquis J, Magistretti J, Trono D, Sandi C. 2013. “Peripuberty stress leads to abnormal aggression, altered amygdala and orbitofrontal reactivity and increased prefrontal MAOA gene expression” Translational Psychiatry 3, e216
http://www.nature.com/tp/journal/v3/n1/full/tp2012144a.html
male rats submitted to fear-induction experiences during the peripubertal period exhibit high and sustained rates of increased aggression at adulthood, even against unthreatening individuals, and increased testosterone/corticosterone ratio. They also exhibit hyperactivity in the amygdala under both basal conditions (evaluated by 2-deoxy-glucose autoradiography) and after a resident–intruder (RI) test (evaluated by c-Fos immunohistochemistry), and hypoactivation of the medial orbitofrontal (MO) cortex after the social challenge. Alterations in the connectivity between the orbitofrontal cortex and the amygdala were linked to the aggressive phenotype
Marsicano G, Moosmann B, Hermann H, Lutz B, Behl C. 2002. “Neuroprotective properties of cannabinoids against oxidative stress: role of the cannabinoid receptor CB1” J Neurochem 80(3): 448-56
http://www.ncbi.nlm.nih.gov/pubmed/11905991/
Neuroprotective effects have been described for many cannabinoids in several neurotoxicity models… CB1 is not involved in the cellular antioxidant neuroprotective effects of cannabinoids
Marsicano G, Wotjak CT, Azad SC, Bisogno T, Rammes G, Cascio MG, Hermann H, Tang J, Hofmann C, Zieglgänsberger W, Di Marzo V, Lutz B. 2002. “The endogenous cannabinoid system controls extinction of aversive memories” Nature 418, 530-534
http://www.nature.com/nature/journal/v418/n6897/abs/nature00839.html
the endogenous cannabinoid system has a central function in extinction of aversive memories. CB1-deficient mice showed strongly impaired short-term and long-term extinction in auditory fear-conditioning tests … endocannabinoids facilitate extinction of aversive memories through their selective inhibitory effects on local inhibitory networks in the amygdala.
Mason M, Norton M, Van Horn J, Wegner D, Grafton S, Macrae C. 2007. “Wandering minds: the default network and stimulus-independent thought” Science 315(5810): 393-395
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1821121/
the mind generally tends to wander, flitting from one thought to the next with fluidity and ease
The default network is minimally disrupted during passive sensory processing and attenuates when people engage in tasks with high central executive demand
We thus trained individuals to become proficient on tasks (10) so that their minds could wander when they performed practiced versus novel task sequences (11)
Despite its regular occurrence, not all minds wander to the same degree; individuals exhibit stable differences in their propensity to produce SIT (1, 3) If regions of the default network underpin the mind’s wandering, then the magnitude of neural activity in these regions should track with people’s proclivity to generate SIT. Specifically, individuals who report frequent mind-wandering should exhibit greater recruitment of the default network when performing tasks that are associated with a high incidence of SIT.
damage to parts of the network (e.g., mPFC) is associated with “mental emptiness” and an absence of spontaneous speech and thought (15) aging is associated with the development of plaques in default network regions and a corresponding reduction in SIT (16, 17)
Reductions in task difficulty are also likely accompanied by qualitative changes in attention and, perhaps, the implementation of general “housekeeping” functions
In light of behavioral evidence suggesting that people are frequently unaware that their mind is wandering (19, 24), it may be the case that the daydream frequency scale (14) used in the current investigation assesses people’s awareness of their mind’s wandering rather than their propensity to engage in SIT.
A second possibility is that SIT—as a kind of spontaneous mental time travel—lends a sense of coherence to one’s past, present, and future experiences (26–29). Finally, the mind may generate SIT not to attain some extrinsic goal (e.g., staying alert) but simply because it evolved a general ability to divide attention and to manage concurrent mental tasks. Although the thoughts the mind produces when wandering are at times useful, such instances do not prove that that the mind wanders because these thoughts are adaptive; on the contrary the mind may wander simply because it can
Mason O, Morgan C, Dhiman S, Patel A, Parti N, Patel A, Curran H. 2009. “Acute cannabis use causes increased psychotomimetic experiences in individuals prone to psychosis” Psychol Med 39(6): 951-6
http://www.ncbi.nlm.nih.gov/pubmed/19017430
A group of cannabis users (n=140) completed the Psychotomimetic States Inventory (PSI) once while acutely intoxicated and again when free of cannabis. A control group (n=144) completed the PSI on two parallel test days. All participants also completed a drug history and the Schizotypal Personality Questionnaire (SPQ). Highly psychosis-prone individuals from both groups were then compared with individuals scoring low on psychosis proneness by taking those in each group scoring above and below the upper and lower quartiles using norms for the SPQ
Smoking cannabis in a naturalistic setting reliably induced marked increases in psychotomimetic symptoms. Consistent with predictions, highly psychosis-prone individuals experienced enhanced psychotomimetic states following acute cannabis use
Maté, G. 2008. In the Realm of Hungry Ghosts: close encounters with addiction. Alfred A Knopf Canada
http://drgabormate.com/book/in-the-realm-of-hungry-ghosts/
Matochik J, Eldreth D, Cadet J, Bolla K. 2005. “Altered brain tissue composition in heavy marijuana users.” Drug Alcohol Depend 77(1):23-30
http://www.ncbi.nlm.nih.gov/pubmed/15607838/ in dropbox
marijuana users had lower gray matter density in a cluster of voxels in the right parahippocampal gyrus, greater density bilaterally near the precentral gyrus and the right thalamus, lower white matter density in the left parietal lobe, and higher density around the parahippocampal and fusiform gyri on the left side compared to non-users
Matsunaga M, Isowa T, Yamakawa K, Fukuyama S, Shinoda J, Yamada J, Ohira H. 2014. “Genetic Variations in the Human Cannabinoid Receptor Gene Are Associated with Happiness” PLoS One 9(4): e93771
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0093771
Mauri M, Volonteri L, De Gaspari I, Colasanti A, Brambilla M, Cerruti L. 2006. “Substance abuse in first-episode schizophrenic patiens: a retrospective study” Clin Pract Epidemiol Ment Health 2: 4
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1435752/
34.7% of first-episode schizophrenic patients had a lifetime history of substance abuse. The age of onset of schizophrenia is significantly lower for drug abusers than for patients without any type of abuse and for alcohol abusers (p < 0.005). In multi drug abusers, cannabis resulted the most frequently used (49%), followed by alcohol (13%), and cocaine (4%)
increases severity of some psychotic symptoms like “hallucination” and “unusual content of thought”
18.10% of the female schizophrenic patients were substance abusers, as against 44.44% of the male patients
substance abusers had significantly higher scores for the “unusual thought content” item (5.5 ± 1.53 vs 5.0 ± 1.51; p < 0.005) and “hostility” item (3.63 ± 2.16 vs 2.64 ± 1.84; p < 0.005), but significantly lower scores for the “depressed mood” item (2.24 ± 1.53 vs 2.71 ± 1.67; p < 0.05)
The age of onset of schizophrenia was significantly lower among the cannabis abusers in comparison with both the cannabis non-abusers (23.97 ± 6.30 vs 26.75 ± 6.45 years; p < 0.05) and the alcohol abusers (23.97 ± 6.30 vs 29.84 ± 8.43 years; p < 0.05) The mean age at first admission of cannabis abusers was significantly lower than that of the non-abusers (25.47 ± 6.10 vs 27.98 ± 6.31 years; p < 0.005), whereas the mean age at first admission of the alcohol abusers was significantly higher than that of both the cannabis non-abusers (32.46 ± 8.05 vs 27.98 ± 6.31 years; p < 0.005) and cannabis abusers (32.46 ± 8.05 vs 25.47 ± 6.10 years; p < 0.005)
The mean “depressed mood” scores tended to be lower among cannabis abusers (1.97 ± 1.27) than among multi-drug abusers (2.55 ± 1.79), non-abusers (2.70 ± 1.67), alcohol abusers (2.15 ± 1.51) and cocaine abusers (3.0 ± 1.82) (p ≤ 0.05)
no statistical difference between the patients with and without a history of substance abuse in terms of a family history of psychiatric disorders
mean duration of hospitalisation was not statistically different between the substance abusers and non-abusers, but was significantly longer among the multi-drug abusers than the single drug abusers (16.03 ± 11.65 vs 23.05 ± 12.34 days; p = 0.006)
possible that substance use improves some latent symptoms experienced by the patients before the onset of schizophrenia [5,11,12]
Mazure C. 1998. “Life stressors as risk factors in depression” Clinical Psychology 5(3): 291-313
http://onlinelibrary.wiley.com/doi/10.1111/j.1468-2850.1998.tb00151.x/abstract
82% of depressive episodes appear to be triggered by major life events
McArdle S. 2010. “Polysubstance Use and Personality Disorder” chapter 9 in Dual Diagnosis: Practice in Context, Blackwell Publishing Ltd. ISBN: 978-1-405-18009-2
http://onlinelibrary.wiley.com/doi/10.1002/9781444314571.ch9/summary in dropbox
they’re associated.
McDonald J, Schleifer L, Richards J, de Wit H. 2003. “Effects of THC on behavioral measures of impulsivity in humans” Neuropsychopharmacology 28: 1356-1365
http://www.nature.com/npp/journal/v28/n7/full/1300176a.html
37 healthy men and women participated in three sessions, in which they received capsules containing placebo, 7.5, or 15 mg THC in randomized order under double-blind conditions
the Stop task, which measures the ability to inhibit a prepotent motor response; a Go/no-go task; a Delay discounting task, which measures the value of delayed or uncertain reinforcers; and a time estimation task, which measures alterations in time perception through a time reproduction procedure.
THC increased impulsive responding on the Stop task but did not affect performance on either the Go/no-go or Delay or Probability discounting tasks. On the time reproduction task, THC increased estimates of the duration of short intervals while not affecting estimates of longer intervals. There were no significant correlations between the four tasks either before or after drug administration
THC was found to increase impulsive responding on the Stop task while not meaningfully affecting performance on the other three measures of impulsive behavior
To say that drugs of abuse, such as marijuana, affect impulsivity, when impulsivity is taken as a nonspecific construct, may not give an accurate or meaningful description of the drug’s effects
McEwen B. 1999. “Stress and hippocampal plasticity” Annu Rev Neurosci 22:105-22
http://www.ncbi.nlm.nih.gov/pubmed/10202533
Repeated stress causes atrophy of dendrites in the CA3 region, and both acute and chronic stress suppresses neurogenesis of dentate gyrus granule neurons. Besides glucocorticoids, excitatory amino acids and N-methyl-D-aspartate (NMDA) receptors are involved in these two forms of plasticity as well as in neuronal death that is caused in pyramidal neurons by seizures and by ischemia
McGee R, Williams S, Poulton R, Moffitt T. 2000. “A longitudinal study of cannabis use and mental health from adolescence to early adulthood” Addiction 95(4): 491-503
http://www.ncbi.nlm.nih.gov/pubmed/10829326
low socioeconomic status in childhood and low parental attachment in adolescence were associated with cannabis use by age 15
McGowan P, Sasaki A, D’Alessio A, Dymov S, Labonte B, Szyf M, Turecki G, Meaney M. 2009. “Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse” Nature Neuroscience 12 342-348
http://www.nature.com/neuro/journal/v12/n3/abs/nn.2270.html
childhood abuse alters HPA stress responses and increases the risk of suicide… decreased levels of glucocorticoid receptor mRNA/mRNA transcripts for glucocorticoid receptor 1F splice variant, increased cytosine methylation of NR3C1 promoter
McLaughlin R, Hill M, Gorzalka B. 2009. “Monoaminergic neurotransmission contributes to cannabinoid-induced activation of the hypothalamic-pituitary-adrenal axis” Eur J Pharmacol 624(1-3): 71-6
http://www.ncbi.nlm.nih.gov/pubmed/19818759/
Administration of high doses of cannabinoid CB(1) receptor agonists activates the hypothalamic-pituitary-adrenal (HPA) axis; however, the mechanism by which this occurs has not been well characterized. Both monoaminergic and glutamatergic neurotransmission are known to activate the HPA axis and cannabinoids have been found to modify levels of these neurotransmitters
acute administration of exogenous cannabinoid ligands activates the HPA axis indirectly through an increase in serotonergic and noradrenergic neurotransmission
McLeod T, Lopez-Figueroa A, Lopez-Figueroa M. 2001. “Nitric oxide, stress, and depression” Psychopharmacol Bull 35(1): 24-41
http://www.ncbi.nlm.nih.gov/pubmed/12397868/
The commonly known neuronal NOS subtype is localized in areas of the brain related to stress and depression. The limbic-hypothalamic-pituitary-adrenal (LHPA) axis is the core of this system. These interrelated pathways have in common the production, and negative feedback, of glucocorticoids. Within these areas, NO is suggested to play a role in modulating the release of other neurotransmitters, acting as a cellular communicator in plasticity and development, and/or acting as a vasodilator in regulation of blood flow
McPartland J, Guy G, Di Marzo V. 2014. “Care and feeding of the endocannabinoid system: a systematic review of potential clinical interventions that upregulate the endocannabinoid system” PLOS ONE 9(3) e89566
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3951193/ in dropbox
Fundamentally, THC mimics AEA and 2-AG by acting as an agonist at CB1 and CB2 [279]. But rather than simply substituting for AEA and 2-AG, McPartland and Guy [280] proposed that Cannabis and its many constituents work, in part, by ‘‘kick-starting’’ the eCB system. The acute administration of THC increased CB1 density in rodent brains [281,282]. Acute upregulation of CB1 mRNA continued for up to 14 days in some rat brain regions [283]. Acute THC also increased the sensitivity of CB1 to cannabinoids, measured by WIN-55,212-2-stimulated [35S]GTP cS binding in rat brains [284]. Lastly, acute THC stimulated AEA biosynthesis [285]
McQueeny T, Padula C, Price J, Medina K, Logan P, Tapert S. 2011. “Gender effects on amygdala morphometry in adolescent marijuana users” Behav Brain Res 224(1): 128-134
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3139567/
“Female marijuana users had larger right amygdala volumes and more internalizing symptoms than female controls, after covarying head size, alcohol, nicotine and other substance use, while male users had similar volumes as male controls. For female controls and males, worse mood/anxiety was linked to smaller right amygdala volume, more internalizing problems was associated with bigger right amygdala in female marijuana users”
… No differences in amygdala size were observed in marijuana using teens compared to non-using peers. … being a marijuana user was associated with worse mood and anxiety symptoms
exclusions for psychiatric diagnosis; mj more alc, nicotine, other drugs; Despite no group-level differences in amygdala volumes, being a marijuana user was associated with worse mood and anxiety symptoms
Mechoulam R, Lichtman A. 2003. “Neuroscience. Stout guards of the central nervous system” Science 302(5642): 65-7
http://www.ncbi.nlm.nih.gov/pubmed/14526067/
activation of the cannabinoid receptor CB1 by the endocannabinoid anandamide protects against excitotoxic damage
Medina K, Schweinsburg A, Cohen-Zion M, Nagel B, Tapert S. 2006. “Effects of alcohol and combined marijuana and alcohol use during adolescence on hippocampal volume and assymetry” Neurotoxicol Teratol 29(1): 141-152
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1821342/
Increased alcohol abuse/dependence severity associated with increased right > left (R>L) asymmetry and smaller left hippocampal volumes; marijuana abuse/dependence was associated with increased L>R asymmetry, larger left hippocampus
smaller left hippocampal volumes among alcohol users… R>L asymmetry was significantly associated with alcohol abuse/dependence severity
R>L asymmetry was associated with improved verbal learning only among the non-substance using control adolescents in this sample
MJ+ALC users did not differ from controls in hippocampal asymmetry or volume … One possibility is that marijuana use could be neuroprotective in combination with alcohol use
Medina K, Nagel B, Park A, McQueeny T, Tapert S. 2008. “Depressive symptoms in adolescents: associations with white matter volume and marijuana use” J Child Psychol Psychiatry 48(6): 592-600
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2269707/ in dropbox
Marijuana and white matter volume additive and interactive in predicting depressive symptoms Subtle neurodevelopmental white matter abnormalities may disrupt the connections between areas involved in mood regulation.
MJ-users and controls did not significantly differ in their intracranial volume (ICV) white matter volume left hippocampal volume or right hippocampal volume
On the HAM-D, 6% of controls and 13% of MJ-users evidenced mild symptoms (scores 7–17) and 6% of MJ-users reported moderate (>18) depressive symptoms
observed interaction between marijuana use and white matter could be due to other moderating factors, such as shared genetic or environmental vulnerability for both disorders
Medina K, Nagel B, Tapert S. 2010. “Abnormal cerebellar morphometry in abstinent adolescent marijuana users.” Psychiatry Res 182(2): 152-9
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2866789/
16 MJ users 1 month abstinent; significantly larger vermis, associated with poorer executive function
non-human animals have greater cerebellar CB1 density than humans (McPartland et al., 2007; Herkenham et al., 1990)
Meier MH, Caspi A, Ambler A, Harrington H, Houts R, Keefe RSE, McDonald K, Ward A, Poulton R, Moffitt T. 2012. “Persistent cannabis users show neuropsychological decline from childhood to midlife.” PNAS 109(40):E2657-E2664
http://www.pnas.org/content/109/40/E2657.full
“ruling out potential confounding effects of (i) acute or residual cannabis intoxication, (ii) tobacco dependence, (iii) hard-drug dependence (e.g., heroin, cocaine, amphetamines), (iv) alcohol dependence, and (v) schizophrenia.”
persistent cannabis dependence showed greater IQ decline. study members who never used cannabis experienced a slight increase in IQ; those diagnosed with cannabis dependence at 1, 2, 3 or more study waves experienced IQ declines of −0.11, −0.17, and −0.38 SD units. An IQ decline of −0.38 SD units corresponds to a loss of ∼6 IQ points, from 99.68 to 93.93. adult-onset cannabis users did not appear to experience IQ decline as a function of persistent cannabis use.
within-person IQ decline was apparent regardless of whether cannabis was used infrequently (median use = 14 d) or frequently (median use = 365 d) in the year before testing.
the impairment was global and detectable across five domains of neuropsychological functioning. impairment was still evident after cessation of use for 1 y or more
there may be some unknown “third” variable that could account for the findings.
the most persistent adolescent-onset cannabis users evidenced an average 8-point IQ decline from childhood to adulthood
Melendez R, Gregory M, Bardo M, Kalivas P. 2004. “Impoverished rearing environment alters metabotropic glutamate receptor expression and function in the prefrontal cortex” Neuropsychopharmacology 29(11): 1980-7
http://www.ncbi.nlm.nih.gov/pubmed/15187985
Rearing rats in impoverished (IC) and enriched (EC) environmental conditions alters synaptic plasticity and cognitive processes. Metabotropic glutamate receptors (mGluRs) are known to play a key role in synaptic and behavioral plasticity… A significant difference in the content of prefrontal mGluR1 and mGluR5 (ie group I) and mGluR2/3 (ie group II) was observed between IC and EC rats
the capacity of group I and II mGluRs to elevate extracellular glutamate levels was significantly blunted in the PFC of IC rats compared to either EC subjects, or rats reared in normal environmental conditions (ie NIH standards). Group II mGluR receptors regulate performance in a forced T-maze spatial memory task that involves the PFC, and IC rats demonstrated deficits in this task relative to EC rats. These data suggest that reduced mGluR transmission in the PFC produced by impoverished, relative to enriched, rearing environments may contribute to cognitive deficits
Meyer-Lindenberg A, Buckholtz J, Kolachana B, Hariri A, Pezawas L, Blasi G, Wabnitz A, Honea R, Verchinski B, Callicott J, Egan M, Mattay V, Weinberger D. 2006. “Neural mechanisms of genetic risk for impulsivity and violence in humans” PNAS 103(16): 6269-6274
http://www.pnas.org/content/103/16/6269.full
low expression variant, [of MAOA gene] associated with increased risk of violent behavior, predicted pronounced limbic volume reductions and hyperresponsive amygdala during emotional arousal, with diminished reactivity of regulatory prefrontal regions, compared with the high expression allele
low expression allele is also associated with changes in orbitofrontal volume, amygdala and hippocampus hyperreactivity during aversive recall, and impaired cingulate activation during cognitive inhibition
One consistent finding in research on MAOA genotype effects on violence and aggression is the more pronounced impact on males
This finding agrees with current proposals linking brain structures involved in emotional control, such as amygdala and medial prefrontal and orbitofrontal cortices, to the emergence of violent behavior (3, 4).
Lesions of OFC are associated with disinhibition and antisocial behavior (3, 4), and we have recently obtained imaging results showing reciprocal regulation of amygdala by both OFC and cingulate (32), suggesting that OFC provides a layer of control that may be especially important if cingulate function is suggested to be compromised, as here
Miller A, Maletic V, Raison C. 2009. “Inflammation and its discontents: the role of cytokines in the pathophysiology of major depression” Biol Psychiatry 65(9): 732-741
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2680424/
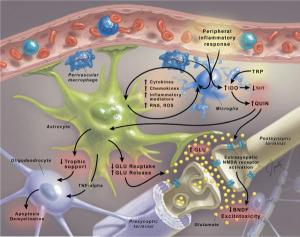
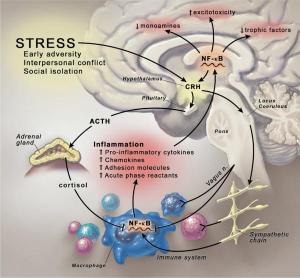
Patients with major depression have been found to exhibit increased peripheral blood inflammatory biomarkers, including inflammatory cytokines, which have been shown to access the brain and interact with virtually every pathophysiologic domain known to be involved in depression, including neurotransmitter metabolism, neuroendocrine function, and neural plasticity
decreased neurotrophic support and altered glutamate release/reuptake, as well as oxidative stress, leading to excitotoxicity and loss of glial elements, consistent with neuropathologic findings that characterize depressive disorders
psychosocial stress, a well-known precipitant of mood disorders, is capable of stimulating inflammatory signaling molecules, including nuclear factor kappa B, in part, through activation of sympathetic nervous system outflow pathways
inhibiting proinflammatory cytokines or their signaling pathways may improve depressed mood and increase treatment response to conventional antidepressant medication
more than 30% of depressed patients fail to achieve remission despite multiple treatment trials (2)
numerous studies have demonstrated a clear relationship between inflammation and the development of cardiovascular disease, diabetes, and cancer (3,4)
dysregulated sleep in depressed patients and sleep deprivation have been associated with increased interleukin (IL)-6, as well as activation of nuclear factor kappa B (NF-κB), a primary transcription factor in the initiation of the inflammatory response (8,10). Although much of the interest in inflammation and depression has been focused on cytokines, which mediate the innate immune response, including IL-1, tumor necrosis factor (TNF)-alpha, and IL-6, which appears to be one of the most reliable peripheral biomarkers in major depression (6,11), findings of increased markers of T cell activation (e.g., soluble IL-2 receptor) in depressed patients raises the specter that both acquired (e.g., T and B cell) and innate (e.g., macrophage) immune responses may be involved (11)
20% to 50% of patients receiving chronic IFN-alpha therapy for the treatment of infectious diseases or cancer develop clinically significant depression (12,13). Of note, depressive syndromes induced by IFN-alpha exhibit considerable overlap with idiopathic major depression and like idiopathic major depression, respond to conventional antidepressant medication (12,13)
cytokine antagonists or anti-inflammatory agents can block the development of behavioral changes following immune activation (17)
binding to cytokine receptors associated with peripheral afferent nerve fibers (e.g., the vagus nerve) that then relay cytokine signals to relevant brain regions including the nucleus of the solitary tract and hypothalamus (17,22)
administration of cytokines or cytokine inducers can profoundly affect the metabolism of serotonin, norepinephrine, and dopamine (DA) (31,32)
Kynurenic acid has been shown to inhibit the release of glutatmate, which, by extension, may inhibit the release of dopamine, whose release is regulated in part by glutamatergic activity (38). Indeed, intrastriatal administration of KA has been shown to dramatically reduce extracellular DA in the rat striatum (39) In contrast, QUIN promotes glutamate release through activation of N-methyl-D-aspartate (NMDA) receptors (). Quinolinic acid also induces oxidative stress, which in combination with glutamate release may contribute to CNS excitotoxicity (see below) (36,40–42). Thus, the relative induction of KA versus QUIN may determine the effects of cytokines on the CNS
Activation of microglia is associated with increased NO production (44), suggesting that cytokine influences on BH4 via NO may be a common mechanism by which cytokines reduce DA availability in relevant brain regions
IL-1 and TNF-alpha have been shown to significantly increase serotonin reuptake in rat brain synaptosomes through activation of p38 MAPK (45)
cytokines may exert a “double hit” on both monoamine synthesis and reuptake, thus contributing to reduced monoamine availability
Cytokines, especially when administered acutely, have been shown to stimulate the expression and release of corticotropin-releasing hormone (CRH) and adrenocorticotropic hormone (ACTH), as well as cortisol, all of which have been found to be elevated in patients with depression (49,50)
impaired negative feedback regulation of HPA axis function is a hallmark of major depression and is reflected by decreased responsiveness to glucocorticoids (or glucocorticoid resistance) as manifested by increased cortisol concentrations following dexamethasone (DEX) administration in the DEX suppression test (DST) and the dexamethasone/corticotrophin-releasing hormone (DEX-CRH) test and decreased glucocorticoid-mediated inhibition of in vitro immune responses (50)
Cytokines can also influence GR expression, leading to decreased GR alpha, the active form of the receptor, and increased GR beta, a relatively inert GR isoform (57)
Cytokines such as IL-1, IL-6, and TNF-alpha that subserve inflammation in the periphery have complex and Janus-faced functional roles in the CNS. Under physiological conditions, these cytokines are important for providing trophic support to neurons and enhancing neurogenesis, while contributing to normal cognitive functions such as memory in laboratory animals (62,63). However, significant data indicate that in the context of excessive and/or prolonged activation, cytokine networks in the CNS can promote an interconnected suite of abnormalities that are increasingly thought to be relevant to the pathophysiology of depression, including diminished neurotrophic support, decreased neurogenesis, increased glutamatergic activation, oxidative stress, induction of apoptosis in relevant cell types (e.g., astrocytes and oligodendrocytes), and dysregulation of glial/neuronal interactions and cognitive function ()(63–77)
acute or chronic stress-leads to increased proinflammatory cytokine production and decreased neurotrophic support and neurogenesis in brain areas important to behavior and cognition (17,64–67)
increased hippocampal concentrations of TNF-alpha and IL-1, which are associated with decreased hippocampal expression of brain-derived neurotrophic factor (BDNF) and its receptor, tyrosine kinase-B, as well as reduced hippocampal neurogenesis (67)
cytokines and inflammatory mediators to increase glutamate release and decrease the expression of glutamate transporters on relevant glial elements, thereby decreasing glutamate reuptake (68,71,75,79, 80)
loss of glial elements, including oligodendrocytes and astrocytes, in multiple mood-relevant brain regions, including the subgenual prefrontal cortex and amygdala, has emerged as a fundamental morphologic abnormality in major depression (74,84,85)
chronic stress, including caregiving, marital discord, and perceived stress, is associated with increases in the acute phase protein, C-reactive protein (CRP), as well as IL-6 and other inflammatory mediators (90–92). In addition, increased inflammation appears to be a hall-mark of early life stress, in that childhood maltreatment has been associated with increased peripheral blood CRP (93). Of note, stress-induced activation of cytokine responses in the CNS appears to be largely dependent on activation of microglia (94)
anti-inflammatory activities of catecholamines have been described (97)
The dACC has been shown to play an important role in error detection and conflict monitoring (104), and increased activity in this brain region has been associated with high trait anxiety, neuroticism, obsessive-compulsive disorder, and bipolar disorder (105)… correlated with task-related emotional distress
Miller G, Chen E, Zhou E. 2007. “If it goes up, must it come down? Chronic stress and the hypothalamic-pituitary-adrenocortical axis in humans” Psychol Bull 133(1):25-45
http://psycnet.apa.org/journals/bul/133/1/25/
hormonal activity is elevated at stressor onset but reduces as time passes. Stressors that threaten physical integrity, involve trauma, and are uncontrollable elicit a high, flat diurnal profile of cortisol secretion. Finally, HPA activity is shaped by a person’s response to the situation; it increases with subjective distress but is lower in persons with posttraumatic stress disorder
Miller G, Rohleder N, Cole S. 2009. “Chronic interpersonal stress predicts activation of pro- and anti- inflammatory signaling pathways six months later” Psychosom Med 71(1): 57-62
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2720615/
To the extent subjects were high in chronic interpersonal stress at baseline, their leukocytes displayed greater increases in mRNA for the pro-inflammatory transcription factor nuclear factor-kappa B (NF-κB) over the next six months. They also showed larger increases in mRNA for inhibitor of kappaB, a molecule that sequesters NF-κB in the cytoplasm and minimizes its pro-inflammatory activities. Chronic interpersonal stress at baseline was unrelated to changes in biomarkers of systemic inflammation, but was associated with increasingly pronounced interleukin-6 responses to lipopolysaccharide. These associations were independent of demographics, lifestyle variables, and depressive symptoms
while this process does not result in systemic inflammation under quiescent conditions, it does accentuate leukocytes’ inflammatory response to microbial challenge. These dynamics may underlie the excess morbidity associated with social stress, particularly in inflammation-sensitive diseases like depression and atherosclerosis
Miller G, Chen E, Fok A, Walker H, Lim A, Nicholls E, Cole S, Kobor M. 2009. “low early-life social class leaves a biological residue manifested by decreased glucocorticoid and increased proinflammatory signaling” Proc Natl Acad Sci U S A 106(34): 14716-14721
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2732821/
Children reared in unfavorable socioeconomic circumstances show increased susceptibility to the chronic diseases of aging when they reach the fifth and sixth decades of life
Among subjects with low early-life SES, there was significant up-regulation of genes bearing response elements for the CREB/ATF family of transcription factors that conveys adrenergic signals to leukocytes, and significant down-regulation of genes with response elements for the glucocorticoid receptor, which regulates the secretion of cortisol and transduces its antiinflammatory actions in the immune system. Subjects from low-SES backgrounds also showed increased output of cortisol in daily life, heightened expression of transcripts bearing response elements for NF-κB, and greater stimulated production of the proinflammatory cytokine interleukin 6. These disparities were independent of subjects’ current SES, lifestyle practices, and perceived stress
suggest that low early-life SES programs a defensive phenotype characterized by resistance to glucocorticoid signaling, which in turn facilitates exaggerated adrenocortical and inflammatory responses
IL-6
Miller B, Buckley P, Seabolt W, Mellor A, Kirkpatrick B. 2011. “Meta-analysis of cytokine alterations in schizophrenia: clinical status and antipsychotic effects” Biol Psychiatry 70(7): 663-671
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4071300/
While some cytokines (IL-1β, IL-6, and TGF-β) may be state markers for acute exacerbations, others (IL-12, IFN-γ, TNF-α, and sIL-2R) may be trait markers
Minzenberg M, Laird A, Thelen S, Carter C, Glahn D. 2009. “Meta-analysis of 41 functional neuroimaging studies of executive function in schizophrenia” Arch Gen Psychiatry 66(8): 811-822
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2888482/
Prefrontal cortical dysfunction is frequently reported in schizophrenia. It remains unclear whether this represents the coincidence of several prefrontal region- and process-specific impairments or a more unitary dysfunction in a superordinate cognitive control network. Whether these impairments are properly considered reflective of hypofrontality vs hyperfrontality remains unresolved
healthy controls and patients activated a similarly distributed cortical-subcortical network, prominently including the dorsolateral prefrontal cortex (PFC), ventrolateral PFC, anterior cingulate cortex (ACC), and thalamus. In between-group analyses, patients showed reduced activation in the left dorsolateral PFC, rostral/dorsal ACC, left thalamus (with significant co-occurrence of these areas), and inferior/ posterior cortical areas. Increased activation was observed in several midline cortical areas
Impaired cognition is a core, disabling feature of schizophrenia, with no established treatment. Although numerous deficits have been described across perceptual, attentional, mnemonic, linguistic, and intellectual functions, impaired executive functions are among the most widely observed, and they are consistently associated with impaired function of the prefrontal cortex (PFC)
A recent quantitative meta-analysis of 12 N-back studies found alterations among schizophrenic subjects compared with healthy controls in several nodes of this neural network, including reduced activity in the DLPFC and increased activity in the VLPFC, ACC, and left frontal pole.2Other studies have reported reduced activity in the VLPFC during other working memory tasks3 and in the ACC during conflict processing
Mitra R, Jadhav S, McEwen B, Vyas A, Chattarji S. 2005. “Stress duration modulates the spatiotemporal patterns of spine formation in the basolateral amygdala” Proc Natl Acad Sci USA 102(26): 9371-6
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1166638/
We find that CIS elicits a robust increase in spine density across primary and secondary branches of BLA spiny neurons… delayed induction of BLA spinogenesis is paralleled by a gradual development of anxiety-like behavior on the elevated plus-maze 10 d after AIS
The basolateral amygdala (BLA) is believed to be a site of storage for memories of fearful or stressful experiences (9–12). Furthermore, recent studies indicate that the synthesis of new proteins in the BLA is involved in the long-term consolidation of emotional memories (13). Thus, the BLA presents an attractive locus to investigate structural encoding of aversive experiences
earlier reports on the role of NMDA receptors in the BLA in fear (19, 20) and anxiety (21)
2 h of immobilization stress per day repeated for 10 consecutive d (CIS), which has previously been reported to cause enhanced anxiety (24, 31) and dendritic growth in the BLA (26, 29), elicits a robust increase in spine density across both primary and secondary branches of BLA spiny neurons
dendritic spines are highly dynamic structures (41–43). During development and even in adulthood, the size and number of spines can be modified by learning, sensory experience, and steroid hormones, as well as many other factors (39, 42, 44–51). For example, there is evidence for greater numbers of synapses per neuron in visual cortex of rats reared in complex environments, and complex motor skill learning leads to an increase in synapse number within the cerebellar cortex (46, 52). Moreover, studies using in vitro techniques have demonstrated that formation of new dendritic protrusions and spines can be triggered by LTP, a synaptic plasticity mechanism thought to underlie learning and memory (4, 53, 54). Such LTP-induced spinogenesis could be mediated by de novospine formation (48) or by splitting of preexisting spines (4, 51, 55, 56)
Lamprecht et al. (15) have demonstrated a role for the Rho-regulatory protein p190 RhoGAP in the lateral amygdala that may contribute to structural changes in synaptic connectivity required for long-term storage of fear memory. This model is particularly interesting because the Rho pathway is known to play a critical role in the developmental regulation of dendritic morphology as well as synaptic connectivity (63–66). Furthermore, there is growing evidence suggesting that the small GTPases of the Rho family might occupy a pivotal role in translating both intra- and extracellular signals into modifications of the actin cytoskeleton (41), which in turn could induce morphological changes of the kind observed after stress.
new spines formed in response to stress may contain synapses with enhanced NMDA conductance, which in turn could enhance calcium influx and affect relevant downstream targets
repeated exposure to the same stressor (CIS) pushes this intracellular machinery to scale up to a greater magnitude of spinogenesis along with enlargement of the dendritic tree
Mobascher A, Winterer G. 2008. “The molecular and cellular neurobiology of nicotine abuse in schizophrenia” Pharmacopsychiatry 41: S51-SF9
https://www.thieme-connect.com/DOI/DOI?10.1055/s-2008-1081463
The effects of nicotine on cognition may be most pertinent to the problem of schizophrenia, but schizophrenics may also smoke to regulate mood and reduce stress
Mokrysz C, Gage S, Landy R, Munafo M, Roiser J, Curran H. 2015. “Neuropsychological and educational outcomes related to adolescent cannabis use, a prospective cohort study” Eur Neuropsychopharmacol 24(2): S695
http://www.ecnp-congress.eu/presentationpdfs/7/P.6.d.016.pdf
ALSPAC cohort, a prospective study following 15,247 pregnancies with expected delivery between April 1991 and December 1992, from Avon, South-West England. Participants completed IQ tests pre-cannabis exposure at age 8 and again at age 15, and completed cannabis-use questionnaires at age 15
Of a complete sample size of 2612, at age 15 years 24a reported trying cannabis at least once. Cannabis use was found to be associated with IQ decline (p≤0.001). Participants who reported using cannabis more than 100 times saw decline of 3.71 IQ points relative to never users (p≤0.010). However once other relevant factors (including sex, socioeconomic status, maternal factors, mental health, and other drug use) were included in the multivariate model the association was attenuated. In particular alcohol use was found to be strongly associated with IQ decline (p≤0.001), and appeared to explain much of the variance in IQ change associated with cannabis use. In those reporting moderate alcohol use (ever use quantity of 6–99 times) this association persisted after control for other relevant factors, however was no longer apparent for heavier alcohol users (ever use quantity of at least 100 times). No other factors were found to be predictive of IQ change. Similar analyses were conducted for educational performance at age 16, controlling for pre-exposure IQ. Cannabis use was no longer associated with educational outcome once controlling for the above factors, however cigarette use remained strongly predictive of educational outcome (p≤0.001).
The findings do not support the hypothesis that cannabis use in adolescence leads to persistent decline in cognitive functioning, once other possible confounding variables are accounted for
Mokrysz C, Landy R, Gage S, Munafo M, Roiser J, Curran H. 2016. “Are IQ and educational outcomes in teenagers related to their cannabis use? A prospective cohort study” Psychopharmacology doi:10.1177/0269881115622241
http://jop.sagepub.com/content/early/2016/01/06/0269881115622241.full
We investigated associations between adolescent cannabis use and IQ and educational attainment in a sample of 2235 teenagers from the Avon Longitudinal Study of Parents and Children. By the age of 15, 24% reported having tried cannabis at least once
After full adjustment, those who had used cannabis ⩾50 times did not differ from never-users on either IQ or educational performance. Adjusting for group differences in cigarette smoking dramatically attenuated the associations between cannabis use and both outcomes, and further analyses demonstrated robust associations between cigarette use and educational outcomes, even with cannabis users excluded. These findings suggest that adolescent cannabis use is not associated with IQ or educational performance once adjustment is made for potential confounds, in particular adolescent cigarette use. Modest cannabis use in teenagers may have less cognitive impact than epidemiological surveys of older cohorts have previously suggested.
Molina-Holgado E, Vela J, Arevalo-Martin A, Almazan G, Molina-Holgado F, Borrell J, Guaza C. 2002. “Cannabinoids promote oligodendrocyte progenitor survival: involvement of cannabinoid receptors and phosphatidylinositol-3 kinase/akt signalling” Journal of Neuroscience 22(22): 9742-9753
http://jneurosci.org/content/22/22/9742.long
Cannabinoids exert pleiotropic actions in the CNS, including the inhibition of inflammatory responses and the enhancement of neuronal survival after injury
Oligodendrocyte progenitors undergo apoptosis with the withdrawal of trophic support, as determined by TUNEL assay and caspase-3 activation, and both the selective CB1 agonist arachidonyl-2′-chloroethylamide/(all Z)-N-(2-cycloethyl)-5,8,11,14-eicosatetraenamide (ACEA) and the nonselective cannabinoid agonists HU210 and (+)-Win-55212-2 enhanced cell survival. To investigate intracellular signaling involved in cannabinoid protection, we focused on the phosphatidylinositol-3 kinase (PI3K)/Akt pathway. HU210, (+)-Win-55212-2, and ACEA elicited a time-dependent phosphorylation of Akt. Pertussis toxin abolished Akt activation, indicating the involvement of Gi/Go-protein-coupled receptors. The CB1 receptor antagonist SR141716A partially inhibited Akt phosphorylation in response to HU210 and (+)-Win-55212-2 and abolished the effects of ACEA. Trophic support deprivation downregulated Akt activity, and cannabinoids recovered phospho-Akt levels. Inhibition of PI3K abrogated the survival action and the recovery of Akt activity in response to cannabinoids. SR141716A prevented only the protection conferred by ACEA. Nevertheless, SR141716A and the selective CB2 receptor antagonist SR144528 in combination inhibited the prosurvival action of HU210, which is in accordance with the finding of CB2 receptor expression by oligodendroglial cells. These data identify oligodendrocytes as potential targets of cannabinoid action in the CNS
Molina P, Amedee A, LeCapitaine N, Zabaleta J, Mohan M, Winsauer P, Stouwe C, McGoey R, Auten M, LaMotte L, Chandra L, Birke L. 2014. “Modulation of Gut-Specific Mechanisms by Chronic Δ9-Tetrahydrocannabinol Administration in Male Rhesus Macaques Infected with Simian Immunodeficiency Virus: A Systems Biology Analysis” AIDS Research and Human Retroviruses 30(6): 567-578
http://online.liebertpub.com/doi/abs/10.1089/aid.2013.0182
Monaghan M, Hamilton I, Lloyd C, Paton K. 2015. “Cannabis matters? Treatment responses to increasing cannabis presentations in addiction services in England” Drugs: Education, Prevention and Policy DOI:10.3109/09687637.2015.1090398
http://www.tandfonline.com/doi/abs/10.3109/09687637.2015.1090398
numerous routes were identified into services for problematic cannabis users
cannabis use was viewed as benign by many staff and clients with noticeable variations of risk
there is an evolving connoisseurship associated with contemporary cannabis use whereby the client has increasing expertise in relation to contemporary cannabis consumption that has yet to fully filter through to the practice of treatment providers… there appears to be a gap between treatment demand and evidence-based treatment for cannabis-related problems
Moore TH, Zammit S, Lingford-Hughes A, Barnes TR, Jones PB, Burke M, Lewis G. 2007. “Cannabis use and risk of psychotic or affective mental health outcomes: a systematic review.” Lancet 370(9584):319-28
http://www.ncbi.nlm.nih.gov/pubmed/17662880 in dropbox
increased risk of any psychotic outcome in ever used cannabis (odds ratio=1.41) greater risk in people who used cannabis most frequently (2.09, 1.54-2.84). … evidence for affective outcomes is less strong… A substantial confounding effect was present for both psychotic and affective outcomes
we conclude that there is now sufficient evidence to warn young people that using cannabis could increase their risk of developing a psychotic illness later in life
Moradi AR, Doost HTN, Taghavi MR, Yule W, Dalgleish T. 2003. “Everyday Memory Deficits in Children and Adolescents with PTSD: Performance on the Rivermead Behavioural Memory Test” Journal of Child Psychology and Psychiatry 40(3):357-361
http://onlinelibrary.wiley.com/doi/10.1111/1469-7610.00453/abstract
Eighteen child and adolescent patients with PTSD and 22 control… PTSD subjects showed poorer overall memory performance (prospective and orientation items of the RBMT)
Morena M, Patel S, Bains J, Hill M. 2015. “Neurobiological interactions between stress and the endocannabinoid system” Neuropsychopharmacology doi:10.1038/npp.2015.166
http://www.nature.com/npp/journal/v41/n1/full/npp2015166a.html
Across a wide array of stress paradigms, studies have generally shown that stress evokes bidirectional changes in the two eCB molecules, anandamide (AEA) and 2-arachidonoyl glycerol (2-AG), with stress exposure reducing AEA levels and increasing 2-AG levels. Additionally, in almost every brain region examined, exposure to chronic stress reliably causes a downregulation or loss of cannabinoid type 1 (CB1) receptors. With respect to the functional role of changes in eCB signaling during stress, studies have demonstrated that the decline in AEA appears to contribute to the manifestation of the stress response, including activation of the hypothalamic–pituitary–adrenal (HPA) axis and increases in anxiety behavior, while the increased 2-AG signaling contributes to termination and adaptation of the HPA axis, as well as potentially contributing to changes in pain perception, memory and synaptic plasticity. More so, translational studies have shown that eCB signaling in humans regulates many of the same domains and appears to be a critical component of stress regulation, and impairments in this system may be involved in the vulnerability to stress-related psychiatric conditions, such as depression and posttraumatic stress disorder
Morgan N, Stanford I, Woodhall G. 2009. “Functional CB2 type cannabinoid receptors at CNS synapses” Neuropharmacology 57(4): 356-68
http://www.ncbi.nlm.nih.gov/pubmed/19616018/
In the medial entorhinal area of the rat, we found that blockade of CB1R did not occlude suppression of GABAergic inhibition by the non-specific endogenous cannabinoid 2-AG… TE-907, a CB2R inverse agonist structurally unrelated to AM-630 elicited increased GABAergic neurotransmission at picomolar concentrations. Analysis of mIPSCs revealed that CB2R effects were restricted to action potential dependent, but not action potential independent GABA release
Moylan S, Maes M, Wray N, Berk M. 2013. “The neuroprogressive nature of mature depressive disorder: pathways to disease evolution and resistance, and therapeutic implications” Mol Psychiatry 18(5): 595-606
http://www.ncbi.nlm.nih.gov/pubmed/22525486
Features of neuroprogression include poorer symptomatic, treatment and functional outcomes in patients with earlier disease onset and increased number and length of depressive episodes. In such patients, longer and more frequent depressive episodes appear to increase vulnerability for further episodes, precipitating an accelerating and progressive illness course leading to functional decline
Murphy J, Houston J, Shevlin M, Adamson G. 2013. “Childhood sexual trauma, cannabis use and psychosis: statistically controlling for pre-trauma psychosis and psychopathology” Soc Psychiatry Psychiatr Epidemiol 48:853-861
http://www.ncbi.nlm.nih.gov/pubmed/23052424 in dropbox
Childhood experiences of rape (B = 0.15) and an interaction between cannabis use and childhood sexual assault ( B = 0.05) independently contributed to adult psychosis. Cannabis use conferred no independent risk.”
consumption among males greater than among females; however, increase in use following trauma is greater among females than among males; cannabis use conferred no independent risk for psychosis
” the ‘cannabis effect’, traditionally identified in analyses devoid of trauma measurement, may have possibly represented a proxy for other more dominant causal agents (e.g. trauma history/traumatic memory)”
consumption is (i) highly associated with a key source of risk for psychosis but does not influence psychosis itself (ii) that it exacerbates another source of risk or moderates pre-existing risk or (iii) that it influences psychosis post-onset
Murillo-Rodriguez E, Poot-Ake A, Arias-Carrion O, Pacheco-Pantoja E, Fuente-Ortegon A, Arankowsky-Sandoval G. 2011. “The emerging role of the endocannabinoid system in the sleep-wake cycle modulation” Cent Nerv Syst Agents Med Chem 11(3): 189-96
http://www.ncbi.nlm.nih.gov/pubmed/21919868/
Murrough J, Perez A, Pillemer S, Stern J, Parides M, aan het Rot M, Collins K, Mathew S, Charney D, Iosifescu D. 2012. “Rapid and longet-term antidepressant effects of repeated ketamine infusions in treatment resistant major depression” Biological Psychiatry 74 (4): 250-256
http://www.biologicalpsychiatryjournal.com/article/S0006-3223(12)00557-4/abstract
Murrough J, Soleimani L, DeWilde K, Collins K, Lapidus K, Iacoviello B, Lener M, Kautz M, Kim J, Stern J, Price R, Perez A, Brallier J, Rodriguez G, Goodman W, Iosifescu D, Charney D. 2015. “Ketamine for rapid reduction of suicidal ideation: a randomized controlled trial” Psychological Medicine 45(16): 3571-3580
http://journals.cambridge.org/action/displayAbstract?fromPage=online&aid=10017684&fulltextType=RA&fileId=S0033291715001506
support for the safety and tolerability of ketamine as an intervention for SI in patients who are at elevated risk for suicidal behavior
Musty R, Kaback L. 1995. “Relationships between motivation and depression in chronic marijuana users” Life Sci 56(23-24): 2151-8
http://www.ncbi.nlm.nih.gov/pubmed/7776845?dopt=Abstract
Several group comparisons were made: Chronic heavy-users (medians: daily use for 6 years) with and without significant symptoms of depression within the last year were compared with Light users (medians: several times per month for 4.5 years) with and without significant symptoms of depression within the last year. … Scores on Need for Achievement were significantly lower in heavy users with depressive symptoms when compared with all other groups. No effects were found among groups in measures of the Need for Affiliation and the Need for Power. Both light and heavy users with symptoms of depression had significantly lower scores than those without depressive symptoms, on the overall Orientation to Life questionnaire and on each subscale measuring Meaningfulness, Manageability and Comprehensibility. These data suggest that amotivational symptoms observed in heavy marijuana users in treatment are due to depression
Navarrete M, Araque A. 2010. “Endocannabinoids potentiate synaptic transmission through stimulation of astrocytes” Neuron 68(1): 113-26
http://www.sciencedirect.com/science/article/pii/S0896627310006860
endocannabinoids released by hippocampal pyramidal neurons increase the probability of transmitter release at CA3-CA1 synapses. This synaptic potentiation is due to CB1R-induced Ca2+ elevations in astrocytes, which stimulate the release of glutamate that activates presynaptic metabotropic glutamate receptors. While endocannabinoids induce synaptic depression in the stimulated neuron by direct activation of presynaptic CB1Rs, they indirectly lead to synaptic potentiation in relatively more distant neurons by activation of CB1Rs in astrocytes
endocannabinoid signaling is generally accepted to induce the inhibition of neurotransmitter release from presynaptic terminals (Chevaleyre et al., 2006 and Hashimotodani et al., 2007)
bidirectional communication between astrocytes and neurons (Araque et al., 2001, Haydon and Carmignoto, 2006, Nedergaard et al., 2003, Perea et al., 2009 and Volterra and Bezzi, 2002). Astrocytes respond with intracellular Ca2+ elevations to neurotransmitters released by synaptic terminals (Perea and Araque, 2005 and Porter and McCarthy, 1996) and, in turn, modulate neuronal excitability and synaptic transmission by releasing neuroactive substances called gliotransmitters (Volterra and Bezzi, 2002, Araque et al., 1999, Beattie et al., 2002 and Perea and Araque, 2007)
Present results indicate that endocannabinoids may have opposite neuromodulatory effects, depressing synaptic transmission by directly acting on presynaptic CB1Rs or potentiating synaptic transmission by acting on astrocytic CB1Rs that stimulate glutamate release from astrocytes
DSE was favored in synapses located at relatively short distance of the endocannabinoid source because the signaling cascade triggered by CB1R activation prevails over that stimulated by mGluR activation (Figure 7)… due to the dendritic arborization, the modulated synapses may be closer to the stimulated neuron than the respective neuronal soma
the fact that homoneuronal synapses only showed DSE suggests that the endocannabinoid-induced synaptic depression mediated by direct activation of presynaptic CB1Rs is the prevailing mechanism over the astrocyte-induced synaptic potentiation. Consequently, it is not surprising that endocannabinoid-induced synaptic potentiation remained elusive when synaptic transmission was analyzed by massive stimulation of axons or when exogenous agonists were applied
In conclusion, astrocytes potentiate hippocampal synaptic efficacy upon stimulation by endocannabinoids, indicating that they are an integral part of the endocannabinoid system. These results expand our view of endocannabinoid signaling as well as astrocytic function on synaptic physiology, and indicate that astrocytes are involved in relevant phenomena of brain function.
Neumeister A, Normandin M. 2013. “Elevated Brain Cannabinoid CB1 Receptor Availability in Posttraumatic Stress Disorder: A Positron Emission Tomography Study” Molecular Psychiatry 18(9): 1034-1040
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3752332/
Three biomarkers examined collectively—OMAR VT, anandamide, and cortisol—correctly classified nearly 85% of PTSD cases
suggest that abnormal CB1 receptor-mediated anandamide signaling is implicated in the etiology of PTSD, and provide a promising neurobiological model [of] pharmacotherapies for this disorder
fear acquisition, consolidation, extinction … neurobiological basis
“development of the next-generation pharmacotherapies”
hyperconsolidation of traumatic memories in PTSD is driven by a glucocorticoid-hormone-facilitated potentiation of norepinephrine inputs to the basolateral amygdala
found that PTSD is associated with a ubiquitously expressed large magnitude elevation (~20%) in [11C]OMAR VT values, which quantitatively reflects CB1 receptor availability
effect found within and outside of amygdala-hippocampal-cortico-striatal neural circuit implicated in PTSD
animal data: sex differences in CB1R regulation, with stress-related up-regulation of CB1 receptors observed predominantly in female animals18
marijuana51 may possess benefits in PTSD – relieve haunting nightmares and other symptoms. such data do not allow the conclusion that self-medication with cannabis with its primary psychoactive constituent tetrahydrocannabinol should be recommended for the treatment of PTSD, as direct activation of CB1 receptors with plant-derived cannabinoids over an extended period of time leads to down-regulation of CB1 receptors52, 53
if, as our results suggest, women show higher CB1 receptor availability than men already under basal, non-stress conditions, then they may be at increased risk for PTSD when exposed to trauma. This finding may thus provide a neurobiological explanation for why women are at greater risk for developing PTSD following exposure to various types of trauma than men even when sexual trauma—which is more common in women—is excluded61
Nguyen B, Kim D, Bricker S, Bongard F, Neville A, Putnam B, Smith J, Plurad D. 2014. “Effect of marijuana use on outcomes in traumatic brain injury” Am Surg 80(10): 979-83
http://www.ncbi.nlm.nih.gov/pubmed/25264643
Nixon S, Paul R, Phillips M. 1998. “Cognitive efficiency in alcoholics and polysubstance users” Alcohol Clin Exp Res 22(7) 1414-20
http://onlinelibrary.wiley.com/doi/10.1111/j.1530-0277.1998.tb03929.x/abstract;jsessionid=8275B93663534625803504A7740B1D41.f03t02
control subjects and individuals who abused both alcohol and marijuana performed significantly better than the other groups on most tests, incl memory, visual/spacial, problem solving
Nomura D, Morrison B, Blankman J, Long J, Kinsey S, Marcondes M, Ward A, Lichtman A, Conti B, Cravatt B. 2011. “Endocannabinoid hydrolysis generates brain prostaglandins that promote neuroinflammation” Science 334(6057): 809-813
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3249428/
Phospholipase A2 (PLA2) enzymes are considered the primary source of arachidonic acid for cyclooxygenase (COX)-mediated biosynthesis of prostaglandins. Here, we show that a distinct pathway exists in brain, where monoacylglycerol lipase (MAGL) hydrolyzes the endocannabinoid 2-arachidonoylglycerol to generate a major arachidonate precursor pool for neuroinflammatory prostaglandins. MAGL-disrupted animals show neuroprotection in a parkinsonian mouse model
MAGL as a distinct metabolic node that couples endocannabinoid to prostaglandin signaling networks in the nervous system and suggest that inhibition of this enzyme may be a new and potentially safer way to suppress the proinflammatory cascades that underlie neurodegenerative disorders
Inflammation is a hallmark of many neurological disorders, including chronic pain, traumatic brain injury, neurodegenerative diseases, and stroke (1). Prominent among the known pro-inflammatory stimuli in the nervous system are prostaglandins, which are produced by cyclooxygenase enzymes (COX1 and COX2) (2) that are in neurons and glial cells (3)
Nooner K, Linares L, Batinjane J, Kramer R, Silva R, Cloitre M. 2012. “Factors Related to Posttraumatic Stress Disorder in Adolescence” Trauma, Violence and Abuse 13(3): 153-66
http://tva.sagepub.com/content/13/3/153.abstract
adolescents are at greater risk of experiencing trauma than either adults or children, and that the prevalence of PTSD among adolescents is 3–57%
Nunn, K. 2002. “Race, crime and the pool of surplus criminality: or why the ‘war on drugs’ was a ‘war on blacks'” Gender Race & Just 6: 381
https://www.google.ca/url?http://www.antoniocasella.eu/archila/Nunn_2002.pdf
Ochoa S, Usall J, Cobo J, Labad X, Kulkarni J. 2012. “Gender differences in schizophrenia and first-episode psychosis: a comprehensive literature review” Schizophrenia Reserach and Treatment 2012, 916198, 9 p
http://www.hindawi.com/journals/schizort/2012/916198/
the Feighner restrictive criteria found a ratio of female-men 0.41 : 1 and the ICD the ratio female-men is 0.92 : 1. Using standard diagnostic criteria in an incidence population study, a meta-analysis by Aleman et al. [4] confirmed that the men had a higher incidence (ratio 1.42). However, the recent studies of prevalence of schizophrenia in general population did not find gender differences [5, 6]
Some authors have suggested that differences in age of onset appear to depend on the presence or absence of family history, with no differences being found between men and women if they had a family history [17, 18]
Nevertheless, most of the studies [15, 26, 27] found no significant clinical differences in symptoms, [19, 26, 27] which is in line with our team’s findings [28]
Several studies have found gender differences in negative symptoms, showing that in males, they were more severe [21–23]. Moreover, in a sample of 276 people with schizophrenia, Galderisi et al. [10] found that men scored higher in disorganization and negative symptoms. In a large sample of patients with psychosis, Morgan et al. [24] identified a higher prevalence of depressive symptoms and lower prevalence of negative symptoms in women. Higher prevalence of depressive and anxiety symptoms in women had been found in previous studies [3, 25] … Results from the assessment of symptoms in first-episode psychosis are also inconclusive. In a group of patients with schizophrenia admitted for the first time, Szymanski et al. [28] found that women presented more anxiety, illogical thinking, inappropriate affect, and bizarre behavior than men. Cotton et al. [29] found that women presented higher levels of affective symptoms than men. However, no gender differences were found in the study by Barajas et al. [30]
Offringa R, Brohawn K, Staples L, Dubois S, Hughes K, Pfaff D, VanElzakker M, Davis F, Shin L. 2013. “Diminished rostral anterior cingulate cortex activation during trauma-unrelated emotional interference in PTSD” Biol Mood Anxiety Disord 3: 10
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3668994/
“relatively diminished rACC activation in PTSD can be observed in interference tasks involving trauma-unrelated emotional stimuli, indicating a more general functional brain abnormality in this disorder”
PTSD in 9-20% of individuals following a traumatic event
impaired behavioral performance and relatively diminished activation in the rostral anterior cingulate cortex (rACC)in Stroop
42 right-handed participants who had been exposed to criterion A traumatic events (e.g., assault, motor vehicle accidents, abuse, and witnessing serious injury/death). 17 PTSD+, 18 PTSD-, no history of drug/alcohol abuse in the last six months
PTSD+ with psychiatric disorders: current major depressive disorder (MDD; n=5), specific phobia (n=1), and panic disorder (n=3)
groups did not significantly differ on response time or error rate measures (ps > 0.32). Only the TENP group exhibited significant rACC activation in the IvC contrast. TENP group exhibited significantly greater rACC activation
significant negative correlation between total CAPS scores and activation of medial frontal gyrus and activation in a more dorsal region of the anterior cingulate cortex
negative correlation between total symptom severity and activation in the rACC proper
Kim et al. [13] found that rACC activation to trauma-unrelated emotional stimuli is negatively correlated with PTSD symptom severity
O’Leary D, Block R, Koeppl J, Flaum M, Schultz S, Andreasen N, Ponto L, Watkins G, Hurtig R, Hichwa R. 2002. “Effects of Smoking Marijuana on Brain Perfusion and Cognition” Neuropsychopharmacology 26 802-816
http://www.nature.com/npp/journal/v26/n6/full/1395868a.html
Smoking marijuana resulted in intoxication, but did not significantly alter mean behavioral performance on the attention task. Heart rate and blood pressure increased dramatically following smoking of marijuana but not placebo
Increased rCBF was observed in orbital and mesial frontal lobes, insula, temporal poles, anterior cingulate, as well as in the cerebellum. The increases in rCBF in anterior brain regions were predominantly in “paralimbic” regions
Reduced rCBF was observed in temporal lobe auditory regions, in visual cortex, and in brain regions that may be part of an attentional network (parietal lobe, frontal lobe and thalamus)
no significant rCBF change in the nucleus accumbens or other reward-related brain regions, nor in basal ganglia or hippocampus, which have a high density of cannabinoid receptors
Oleson E, Cheer J. 2012. “A brain on cannabinoids: the role of dopamine release in reward seeking” Cold Spring Harbor Perspectives in Medicine doi: 10.1101/cshperspect.a012229
http://perspectivesinmedicine.cshlp.org/content/2/8/a012229.full
Increases in mesolimbic dopamine transmission are observed when animals are treated with all known drugs of abuse, including cannabis, and to conditioned stimuli predicting their availability… despite general misconceptions that cannabis is unique from other drugs of abuse, cannabis exerts identical effects on the mesolimbic dopamine system
[MV: study is based entirely on rodent findings. Studies in humans have failed to find comparable results. The fact that cannabis has identical effects on mesolimbic dopamine signalling to other drugs of abuse in rats, but not in humans, casts suspicion on the relevance of murine models to human cannabis use]
Onaivi E, Ishiguro H, Gong J-P, Patel S, Meozzi P, Myers L, Perchuk A, Mora Z, Tagliaferro P, Gardner E, Brusco A, Akinshola B, Liu Q-R, Chirwa S, Hope B, Lujilde J, Inada T, Iwasaki S, Macharia D, Teasenfitz L, Arinami T, Uhl G. 2008. “Functional expression of brain neuronal CB2 cannabinoid receptors are involved in the effects of drugs of abuse and in depression” Ann N Y Acad Sci 1139: 434-449
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3922202/
high incidence of (Q63R) but not (H316Y) polymorphism in the CB2 gene was found in Japanese depressed subjects
Mice that developed alcohol preference had reduced CB2 gene expression and chronic treatment with JWH015 a putative CB2-R agonist, enhanced alcohol consumption in stressed but not in control mice. The direct intracerebroventricular microinjection of CB2 anti-sense oligonucleotide into the mouse brain reduced mouse aversions in the plus-maze test, indicating the functional presence of CB2-Rs in the brain that modifies behavior
de Oliveira Alvares L, Pasqualini Genro G, Diehl F, Molina V, Quillfeldt J. 2008. “Opposite action of hippocampal CB1 receptors in memory reconsolidation and extinction” Neuroscience 154(4): 1648-55
http://www.ncbi.nlm.nih.gov/pubmed/18554811/
reconsolidation requires a brief reactivation session, and extinction, a prolonged one
Local infusion of AM251 into CA1 blocked memory extinction whereas the administration of anandamide facilitated it; however, when combined with a subthreshold concentration of the CB1 antagonist, anandamide did not affect the extinction process. The clear-cut, opposite effects observed in each situation suggest a possible role of the hippocampal endocannabinoid system as a switching mechanism deciding which processes will take place, either maintaining the original memory (reconsolidation) or promoting a new learning (extinction)
van Os J, Bak M, Hanssen M, Bijl R, de Graaf R, Verdoux H. 2002. “Cannabis Use and Psychosis: A Longitudinal Population-based Study.” Am J Epidemiol 156(4): 319-327
http://aje.oxfordjournals.org/content/156/4/319.long
Baseline cannabis use predicted the presence at follow-up of any level of psychotic symptoms (adjusted odds ratio (OR) = 2.76
van Os J, Linscott R, Myin-Germeys I, Delespaul P, Krabbendam L. 2009. “A systematic review and meta-analysis of the psychosis continuum: evidence for a psychosis proneness-persistence-impairment model of psychotic disorder” Psychological Medicine 39(2): 179-195
http://journals.cambridge.org/action/displayAbstract?fromPage=online&aid=3405404&fileId=S0033291708003814
A systematic review of all reported incidence and prevalence studies of population rates of subclinical psychotic experiences reveals a median prevalence rate of around 5% and a median incidence rate of around 3%. A meta-analysis of risk factors reveals associations with developmental stage, child and adult social adversity, psychoactive drug use, and also male sex and migrant status. The small difference between prevalence and incidence rates, together with data from follow-up studies, indicates that approximately 75–90% of developmental psychotic experiences are transitory and disappear over time
he psychosis proneness–persistence–impairment model considers genetic background factors impacting on a broadly distributed and transitory population expression of psychosis during development, poor prognosis of which, in terms of persistence and clinical need, is predicted by environmental exposure interacting with genetic risk
Otte C, Neylan T, Pole N, Metzler T, Best S, Henn-Haase C, Yehuda R, Marmar C. 2005. “Association between childhood trauma and catecholamine response to psychological stress in police academy recruits” Biol Psychiatry 57: 27-32
http://www.ncbi.nlm.nih.gov/pubmed/15607297 in dropbox
6 police academy recruits (mean [ SD] age 28 5 years, 10 female) were exposed to a video depicting real-life officers exposed to highly stressful incidents. Salivary cortisol and 3-methoxy-4-hydroxy-phenylglycol (MHPG, the major metabolite of norepinephrine) were collected at baseline, immediately after the video, and 20 min after the video. Childhood trauma before age 14 was assessed with an interview (Life Stressor Checklist–Revised)
Recruits with childhood trauma histories (n 16) had a significantly greater MHPG response, as evidenced by a group effect (F 8.0, p .01), and a group time interaction (F 4.1, p .05). The cortisol response did not differ between groups.
In the United States, more than 2.5 million cases of child abuse and neglect are reported each year (American Acad-emy of Pediatrics 2004)
one possible explanation for our results is that trauma exposure during childhood, a period of heightened plasticity of corticolimbic neuronal systems, in-creases amygdala reactivity in adulthood to novel stressful or even neutral situations, so that subsequent responses are affected
Childhood trauma might lead to a lower threshold for interpreting stimuli as threatening and to a selective attention bias toward searching for and identifying threatening information, leading to a higher catechol-amine response (Foa et al 1989)
Greater salivary MHPG during memory consolidation was associated with greater recall for an emotionally arousing story 1 week later (Southwick et al 2002). Importantly, it has been shown that salivary MHPG strongly correlates with MHPG in the cerebrospinal fluid (Reuster et al 2002), suggesting that salivary MHPG is a reliable proxy of brain MHPG activity
only maladapted, behaviorally “symptomatic” animals, but not well-adapted or control rats, show an increased cortisol response to stress (Cohen et al 2003)
we did not measure ACTH and cannot exclude the possibility that there were differences in pituitary responsiveness between groups
Pace T, Hu F, Miller A. 2007. “Cytokine-effects on glucocorticoid receptor function: relevance to glucocorticoid resistance and the pathophysiology and treatment of major depression” Brain Behav Immun 21(1): 9-19
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1820632/
inflammatory cytokines and their signaling pathways including mitogen-activated protein kinases, nuclear factor-kB, signal transducers and activators of transcription, and cyclooxygenase have been found to inhibit GR function. Mechanisms include disruption of GR translocation and/or GR-DNA binding through protein-protein interactions of inflammatory mediators with the GR itself or relevant steroid receptor cofactors as well as alterations in GR phosphorylation status. Interestingly, cAMP signal transduction pathways can enhance GR function and inhibit cytokine signaling
a number of disorders characterized by excessive inflammatory responses including rheumatoid arthritis, asthma, and inflammatory bowel disease as well as depression have been associated with resistance to the inhibitory effects of glucocorticoids (Pariante and Miller, 2001; Raison and Miller, 2003). In the case of major depression, a clinical disorder characterized by significant alterations in mood, neurovegetative function and cognition, glucocorticoid resistance has been one of the most reproducible biological findings in the disease, occurring in up to 80% of patients (see below) (Holsboer, 2000; Pariante and Miller, 2001; Heuser et al., 1994)
One of the most reliably reported neurobiological alterations in major depression is both HPA axis hyperactivity and impaired HPA axis glucocorticoid feedback sensitivity
CRH hyperactivity in major depression is believed to be related, in part, to the failure of cortisol to suppress CRH production through negative feedback regulation (Holsboer, 2000; Pariante and Miller, 2001)
major depression is associated with immune activation as reflected by increased plasma and CSF concentrations of a variety of cytokines and their receptors, including IL-1, IL-2, IL-6 and TNF-alpha, as well as increases in plasma concentrations of acute phase reactants (which reflect the effects of proinflammatory cytokines on the liver), chemokines, and cellular adhesion molecules (Raison et al., 2006) Probably the most reproducible finding of immune activation in patients with depression is increased plasma levels of IL-6 and its downstream product from the liver, C-reactive protein (CRP).
cytokine-induced increases in dACC activity may reflect an increased sensitivity to error (and/or negatively perceived internal or external events) and thus potentially represent a cognitive pathway to psychopathology during cytokine exposure
in patients with major depression, increased mitogen-induced IL-1 responses of peripheral blood mononuclear cells (PBMCs) were found to positively correlate with post-DST plasma levels of cortisol, suggesting that DST non-suppression may be related to the effects of pro-inflammatory cytokines on GR signaling (Maes et al., 1993)
within the cell, GR reside primarily in the cytoplasm, stabilized in the non-activated state by association with a chaperone protein complex containing heat shock proteins. Upon ligand binding and activation, the GR dissociates from the chaperone protein complex, becomes hyperphosphorylated, undergoes a conformational change and translocates to the nucleus. Once in the nucleus dimerization occurs, and GR homodimers influence cellular function through either interaction with other nuclear transcription factors or binding to the glucocorticoid responsive promoter region (glucocorticoid responsive element; GRE) of relevant genes. GR-DNA binding requires a number of steroid receptor co-factors for initiation of promoter activity. Protein-protein interactions of the GR with NF-kB and GR-GRE binding leading to the induction of IkB (which stabilizes non-activated NF-kB in the cytosol) are major mechanisms by which the GR regulates inflammatory responses (McKay and Cidlowski, 1999). … protein-protein interactions (GR-NF-kB) may be a more primary contributor to glucocorticoid inhibition of NF-kB than induction of IkB. In addition, overexpression of GR results in a marked inhibition of NF-kB activity even in the absence of steroid,(Raddatz et al., 2001) further supporting the notion that NF-kB-GR interactions play an important role in balancing responses to stress and inflammation
Pagliaccio D, Barch D, Bogdan R, Wood P, Lynskey M, Heath A, Agrawal A. 2015. “Shared predisposition in the association between cannabis use and subcortical brain structure” JAMA Psychiatry 72(1): 994-1001
http://www.ncbi.nlm.nih.gov/pubmed/26308883
Among 483 study participants, cannabis exposure was related to smaller left amygdala (approximately 2.3%; P = .007) and right ventral striatum (approximately 3.5%; P < .005) volumes. These volumetric differences were within the range of normal variation. The association between left amygdala volume and cannabis use was largely owing to shared genetic factors (ρg = -0.43; P = .004), while the origin of the association with right ventral striatum volumes was unclear. Importantly, brain volumes did not differ between sex-matched siblings discordant for use (fixed effect = -7.43; t = -0.93, P = .35). Both the exposed and unexposed siblings in pairs discordant for cannabis exposure showed reduced amygdala volumes relative to members of concordant unexposed pairs (fixed effect = 12.56; t = 2.97; P = .003)
we found no evidence for the causal influence of cannabis exposure on amygdala volume
Palamar J, Fenstermaker M, Kamboukos D, Ompad D, Cleland C, Weitzman M. 2014. “Adverse psychosocial outcomes associated with drug use among US high school seniors: a comparison of alcohol and marijuana” Am J Drug Alcohol Abuse 1-9
http://informahealthcare.com/doi/abs/10.3109/00952990.2014.943371
marijuana use was more commonly reported to compromise relationships with teachers or supervisors, result in less energy or interest, and result in lower school or job performance. Compared to marijuana use, alcohol was more commonly reported to compromise relationships with friends and significant others; it was also reported to lead to more regret (particularly among females), and driving unsafely. Marijuana users were more likely to report no adverse outcomes
some adverse outcomes experienced by marijuana users are likely related to marijuana policy rather than use per se
Palazuelos J, Aguado T, Pazos M, Julien B, Carrasco C, Resel E, Sagredo O, Benito C, Romero J, Azcoitia I, Fernandez-Ruiz J, Guzman M, Galve-Roperh I. 2009. “Microglial CB2 cannabinoid receptors are neuroprotective in Huntington’s disease excitotoxicity” Brain 132(Pt 11): 3152-64
http://www.ncbi.nlm.nih.gov/pubmed/19805493
Cannabinoid-derived drugs are promising agents for the development of novel neuroprotective strategies. Activation of neuronal CB(1) cannabinoid receptors attenuates excitotoxic glutamatergic neurotransmission, triggers prosurvival signalling pathways and palliates motor symptoms in animal models of neurodegenerative disorders
we show that CB(2) cannabinoid receptor expression increases in striatal microglia of Huntington’s disease transgenic mouse models and patients. Genetic ablation of CB(2) receptors in R6/2 mice, that express human mutant huntingtin exon 1, enhanced microglial activation, aggravated disease symptomatology and reduced mice lifespan. Likewise, induction of striatal excitotoxicity in CB(2) receptor-deficient mice by quinolinic acid administration exacerbated brain oedema, microglial activation, proinflammatory-mediator state and medium-sized spiny neuron degeneration. Moreover, administration of CB(2) receptor-selective agonists to wild-type mice subjected to excitotoxicity reduced neuroinflammation, brain oedema, striatal neuronal loss and motor symptoms
a pivotal role for CB(2) receptors in attenuating microglial activation and preventing neurodegeneration that may pave the way to new therapeutic strategies for neuroprotection in Huntington’s disease as well as in other neurodegenerative disorders with a significant excitotoxic component
Pamplona F, Bitencourt R, Takahashi R. 2008. “Short-and long-term effects of cannabinoids on the extinction of contextual fear memory in rats” Neurobiol Learn Mem 90(1): 290-3
http://www.ncbi.nlm.nih.gov/pubmed/18486502
CB1 agonist injection boosts contextual fear extinction in rats
Panikashvili D, Shein N, Mechoulam R, Trembovler V, Kohen R, Alexandrovich A, Shohami E. 2006. “The endocannabinoid 2-AG protects the blood-brain barrier after closed head injury and inhibits mRNA expression of proinflammatory cytokines” Neurobiol Dis 22(2): 257-64
http://www.ncbi.nlm.nih.gov/pubmed/16364651/
the endocannabinoid 2-arachidonoyl glycerol (2-AG) is released in mouse brain after closed head injury (CHI), and treatment with exogenous 2-AG exerts neuroprotection via the central cannabinoid receptor CB1. This process involves inhibition of inflammatory signals that are mediated by activation of the transcription factor NF-kB… We found that 2-AG decreased BBB permeability and inhibited the acute expression of the main proinflammatory cytokines: TNF-alpha, IL-1beta and IL-6. It also augmented the levels of endogenous antioxidants. We suggest that 2-AG exerts neuroprotection in part by inhibition of the early (1-4 h) inflammatory response and augmentation of the brain reducing power
Passie T, Emrich H, Karst M, Brandt S, Halpern J. 2012. “Mitigation of post-traumatic stress symptoms by Cannabis resin: a review of the clinical and neurobiological evidence” Drug Test Anal 4(7-8):649-59
http://www.ncbi.nlm.nih.gov/pubmed/22736575
Case report of 19 year old for whom cannabis resin dramatically reduced severe PTSD symptoms.
Cannabis may dampen the strength or emotional impact of traumatic memories through synergistic mechanisms that might make it easier for people with PTSD to rest or sleep and to feel less anxious and less involved with flashback memories
Pariante C. 2003. “Depression, stress and the adrenal axis” J Neuroendocrinology 15(8):811-2
http://www.ncbi.nlm.nih.gov/pubmed/12834443
Depression is characterized by an over activity of the hypothalamic-pituitary-adrenal (HPA) axis
Parker LA, Burton P, Sorge RE, Yakiwchuk C, Mechoulam R. 2004. “Effect of low doses of delta9-tetrahydrocannabinol and cannabidiol on the extinction of cocaine-induced and amphetamine-induced conditioned place preference learning in rats.” Psychopharmaceology (Berl) 175(3):360-6
http://www.ncbi.nlm.nih.gov/pubmed/15138755
it accelerates it (easier to unlearn conditioned associations)
Patel S, Roelke CT, Rademacher DJ, Cullinan WE, Hillard CJ. 2004. “Endocannabinoid signalling negatively modulates stress-induced activation of the hypothalamic-pituitary-adrenal axis.” Endocrinology 145(12):5431-8
http://www.ncbi.nlm.nih.gov/pubmed/15331569
eCB signaling negatively modulates HPA axis function in a context-dependent manner and suggest that pharmacological augmentation of eCB signaling could serve as a novel approach to the treatment of anxiety-related disorders.
Patel S, Hillard C. 2009. “Role of Endocannabinoid Signalling in Anxiety and Depression” Current Topics in Behavioural Neuroscience
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3808114/
Patton G, Coffey C, Carlin J, Degenhardt L, Lynskey M, Hall W. 2002. “Cannabis use and mental health in young people: cohort study” BMJ 325
http://www.bmj.com/content/325/7374/1195.1
daily use in female teenagers predicted fourfold higher odds of later depression and anxiety (odds ratio 4.2, 1.6 to 11), weekly use a twofold elevation (2.3, 1.3 to 4.2)
The association with cannabis use persisted after adjustment for concurrent use of alcohol, tobacco, and other illicit substances as well as indices of family disadvantage
Pava M, Woodward J. 2012. “A Review of the Interactions between Alcohol and the Endocannabinoid System: Implications for Alcohol Dependence and Future Directions for Research” Alcohol 46(3): 185-204
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3327810/
Alcohol use disorders (AUDs) affect an estimated 8.5% of the US population over the age of 18… cost the United States economy up to $185 billion per year
Marijuana users have cross tolerance to alcohol; 45-81% of CUD have AUD; 70% of adolescents with AUD used cannabis in last year
cannabis users significantly increased ethanol craving/consumption during abstinence from marijuana (Peters and Hughes, 2010) treatment for CUDs increased frequency of ethanol use during 12 months following treatment (Stephens et al., 1994)
Jones and Stone (1970) heavy marijuana use were less intoxicated by ethanol and showed fewer ethanol-induced neuropsychological impairments than were found in previous studies
the functional consequences of CB1 activation are determined by the ligand (distinct signal transduction cascades from different ligands)
CB1R decreased in mice chronically exposed to ethanol
EC synthesis/release dependent on excitability induced elevations in intracellular Ca2+; brief incubation with ethanol is known to increase intracellular Ca2+ in forebrain synaptosomes (Davidson et al., 1988) alcohol thereby increases AEA and 2-AG
rewarding properties of ethanol may be reduced following chronic exposure to cannabinoids
Direct measures of EC tissue content have found that acute or short-term exposure to ethanol reduces EC levels in the hippocampus, striatum, prefrontal cortex, amygdala, and cerebellum (Ferrer et al., 2007; Rubio et al., 2007)
EC release from medium-spiny neurons in the dorsomedial striatum is associated with a long-lasting disinhibition of these cells, and this effect is blocked by pre-treatment with ethanol (50 mM) via a pre-synaptic mechanism that is independent of EC synthesis and CB1 activation (Clarke and Adermark, 2010) activation of CB1 by WIN is sufficient to block pre-synaptic facilitation of GABAergic signaling on pyramidal neurons (PNs) in the central amygdala produced by acutely applied ethanol (Roberto et al., 2010)
future studies should be sensitive to the possible interactions between the EC system and the state of the organism under study
rodents with fewer CB1R have stronger preference for ethanol; more intake leads to higher 2-AG/AEA levels
Pawlak R, Magarinos A, Melchor J, McEwen B, Strickland S. 2003. “Tissue plasminogen activator in the amygdala is critical for stress-induced anxiety-like behaviour” Nat Neurosci 6(2): 168-74
http://www.ncbi.nlm.nih.gov/pubmed/12524546/
serine protease tissue-plasminogen activator (tPA) was upregulated in the central and medial amygdala by acute restraint stress, where it promoted stress-related neuronal remodeling
Pepin M, Pothier F, Barden N. 1992. “Impaired type II glucocorticoid-receptor function in mice bearing antisense RNA transgene” Nature 355(6362): 725-8
http://www.ncbi.nlm.nih.gov/pubmed/1741058/
Glucocorticoids, in conjunction with their cognate receptors, exert negative-feedback effects on the hypothalamus-pituitary-adrenal axis, suppressing adrenal steroid secretions. Two types of corticosteroid receptor, distinguishable by their ability to bind corticosterone, have been identified as classical mineralocorticoid (type I) and glucocorticoid (type II) receptors by cloning their complementary DNAs. The type I receptor controls the basal circadian rhythm of corticosteroid secretion. Both receptor types are involved in negative feedback, but the type II receptor may be more important for terminating the stress response as it is the only one to be increased in animals rendered more sensitive to corticosteroid negative-feedback effects
Peralta V, Cuesta M. 1992. “Influence of cannabis abuse on schizophrenic psychopathology” Acta Psychiatr Scand 85(2): 127-30
http://www.ncbi.nlm.nih.gov/pubmed/1543037
95 schizophrenia patients, of whom 23 were cannabis abusers
no statistically significant differences between groups on scales of assessment of positive symptoms (SAPS) — cannabis uers had higher scores except for the delusions subscale. On the SANS nonabusers scored higher, with a significant difference on the alogia subscale.
consumption of cannabis by schizophrenic patients could attenuate negative symptoms, which would support the self-medication hypothesis of cannabis abuse
Pedersen B, Fischer C. 2007. “Physiological roles of muscle-derived interleukin-6 in response to exercise” Curr Opin Clin Nutr Metab Care 10(3): 265-71
http://www.ncbi.nlm.nih.gov/pubmed/17414493
Contraction-induced transcription and release of interleukin-6 is primarily regulated by an altered intramuscular milieu in response to exercise. Accordingly, changes in calcium homeostasis, impaired glucose availability and increased formation of reactive oxygen species are all associated with exercise and capable of activating transcription factors known to regulate interleukin-6 synthesis. Acute interleukin-6 administration to humans increases lipolysis, fat oxidation and insulin-mediated glucose disposal. Adenosine monophosphate-activated protein kinase activation by interleukin-6 appears to play an important role in modulating some of these metabolic effects. Interleukin-6 facilitates an antiinflammatory milieu and may exert some of its biological effects via inhibition of the proinflammatory cytokine tumor necrosis factor-alpha (TNFα)
Perez C, Widom C. 1994. “Childhood victimization and long-term intellectual and academic outcomes” Child Abuse Negl 18(8): 617-33
http://www.ncbi.nlm.nih.gov/pubmed/7953902/ in dropbox
Abused and neglected and control group individuals differed significantly in IQ and reading ability, even when controlling for age, sex, race, and social class. Types of maltreatment were associated with differences in IQ and reading ability in young adulthood
58% of the abused/neglected group did not complete high school compared to 34% of the controls.
neglected children at approximately age 28 were significantly different from controls on both IQ and WRAT, that physically abused individuals differed only on IQ, and that sexually abused children did not differ from controls in IQ and WRAT reading
Perry J. 1974. The Far Side of Madness Prentice-Hall
http://books.google.ca/books/about/The_far_side_of_madness.html?id=ywRsAAAAMAAJ
Perry B, Pollard R, Blakley T, Baker W, Vigilante D. 1995. “Childhood Trauma, the Neurobiology of Adaptation & Use-dependent Development of the Brain: How States become Traits” Infant Mental Health Journal
http://www.trauma-pages.com/a/perry96.php
at the time when the human is most vulnerable to the effects of trauma — during infancy and childhood — adults generally presume the most resilience.
Neurotransmitter receptor/effector activation then alters intracellular chemical constituents, including second (e.g. cAMP, phosphatidyl inositol) and third messengers (e.g., calcium).
The child who has been emotionally neglected early in life will exhibit profound attachment problems which are extremely insensitive to any replacement experiences later in life, including therapy
discussion of sensitized hyperarousal (more in boys) vs dissociation (more in girls) the younger an individual is, the more likely they are to use dissociative adaptations over hyperarousal responses.
Only 70 percent of our severely traumatized children (all ages) with dramatic symptoms of physiological hyperarousal (n=120; males/females: 4:1) meet diagnostic criteria for PTSD (Perry, 1995). Furthermore, in our experience, females have many more affective (dysthymia, major depression), dissociative (65 % with Child Dissociative Checklist scores over cutoff) and borderline personality features than males
they frequently misunderstand the child’s unattached, nonreactive behaviors as ‘not being effected’ rather than the ‘surrender’ response
Children are not resilient, children are malleable. In the process of getting over it, elements of their true emotional, behavioral, cognitive and social potential are diminished — some percentage of capacity is lost, a piece of the child is lost forever.
Persistence of the destructive myth that ‘children are resilient’ will prevent millions of children, and our society, from meeting their true potential. Persistence of the pervasive maltreatment of children in the face of decreasing global and national resources will lead, inevitably, to sociocultural devolution.
It need not be so.
Pertwee R. 2006. “The pharmacology of cannabinoid receptors and their ligands: an overview” International Journal of Obesity 30 S13-S18
http://www.nature.com/ijo/journal/v30/n1s/full/0803272a.html
For CB1 receptor agonists, other potential uses include the management of glaucoma, pain, certain types of cancer and various kinds of motor dysfunction associated for example with multiple sclerosis or spinal cord injury.8, 12, 22, 23 Particularly convincing are preclinical, anecdotal and clinical data supporting the use of CB1 receptor agonists against inflammatory and neuropathic pain and for the amelioration of spasticity, muscle spasms, tremor or pain associated with multiple sclerosis or spinal cord injury.8, 12 Indeed, a  9-THC-containing extract of cannabis has now been developed in the UK as a medicine for the management of some multiple sclerosis symptoms. As to CB2 receptor agonists, there is already strong evidence that these have potential for the relief of inflammatory pain and, unexpectedly, also for the relief of neuropathic pain.24
9-THC-containing extract of cannabis has now been developed in the UK as a medicine for the management of some multiple sclerosis symptoms. As to CB2 receptor agonists, there is already strong evidence that these have potential for the relief of inflammatory pain and, unexpectedly, also for the relief of neuropathic pain.24
Peterson B, Potenza M, Wang Z, Zhu H, Martin A, Marsh R, Plessen K, Yu, S. 2009. “An fMRI study of the effects of psychostimulants on default-mode processing during stroop task performance in youths with ADHD” Am J Psychiatry 166(11): 1286-1294
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3289412/
Stimulant medication significantly improved suppression of default-mode activity in the ventral anterior cingulate cortex in the ADHD group. When off medication, youths with ADHD were unable to suppress default-mode activity to the same degree as comparison subjects, whereas when on medication, they suppressed this activity to comparison group levels. Greater activation of the lateral prefrontal cortex when off medication predicted a greater reduction in ADHD symptoms when on medication. Granger causality analyses demonstrated that activity in the lateral prefrontal and ventral anterior cingulate cortices mutually influenced one another but that the influence of the ventral anterior cingulate cortex on the lateral prefrontal cortex was significantly reduced in youths with ADHD off medication relative to comparison subjects and increased significantly to normal levels when ADHD youths were on medication
Brain regions that have been shown with fair consistency to activate less in children with ADHD compared with healthy comparison subjects during performance of tasks that require attention and inhibitory control include the striatum (1–3), the anterior cingulate cortex (4), the prefrontal cortex (3, 5, 6), and the inferior frontal gyrus (5, 7)
Phan K, Britton J, Taylor S, Fig L, Liberzon I. 2006. “Corticolimbic blood flow during nontraumatic emotional processing in posttraumatic stress disorder” Arch Gen Psychiatry 63(2): 184-92
http://www.ncbi.nlm.nih.gov/pubmed/16461862/
Consistent with current functional neuroanatomic models, patients with PTSD exhibited altered neural responses in the amygdala and ventral MPFC during the processing of emotionally salient but trauma-unrelated stimuli
Phan K, Angstadt M, Golden J, Onyewuenyi I, Popovska A, de Wit H. 2008. “Cannabinoid modulation of amygdala reactivity to social signals of threat in humans” J Neurosci 28(10): 2313-2319
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2657360/
Poirier j, Delisle M, Quirion R. 1995. “Apolipoprotein E4 allele as a predictor of cholinergic deficits and treatment outcome in Alzheimer’s disease” Proc Natl Acad Sci 92: 12260-4
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC40336/
Acetylcholine esterase inhibition may exhibit or lack cognitive benefits depending on genotype
van der Pol P, Liebregts N, de Graaf R, Korf D, van den Brink W, van Laar M. 2013. “Predicting the transition from frequent cannabis use to cannabis dependence: a three-year prospective study” Drug Alcohol Depend 133(2): 352-9
http://www.drugandalcoholdependence.com/article/S0376-8716(13)00228-7/fulltext
A prospective cohort of frequent cannabis users (aged 18-30, n=600) with baseline and two follow-up assessments (18 and 36 months) was used. Only participants without lifetime diagnosis of DSM-IV cannabis dependence at baseline (n=269) were selected
variables assessed as potential predictors of the development of cannabis dependence included sociodemographic factors, cannabis use variables (e.g., motives, consumption habits, cannabis exposure), vulnerability factors (e.g., childhood adversity, family history of mental disorders or substance use problems, personality, mental disorders), and stress factors (e.g., life events, social support)
Three-year cumulative incidence of cannabis dependence was 37.2% (95% CI=30.7-43.8%). Independent predictors of the first incidence of cannabis dependence included: living alone, coping motives for cannabis use, number and type of recent negative life events (major financial problems), and number and type of cannabis use disorder symptoms (impaired control over use). Cannabis exposure variables and stable vulnerability factors did not independently predict first incidence of cannabis dependence
[MV: excluding cannabis dependence at baseline among frequent users probably eliminated the effect of childhood adversity and other vulnerability factors]
The distinction between non-dependent and dependent frequent use is important, because dependent users, by definition, experience significant psychosocial impairments related to reduced control over their cannabis use, whereas other frequent users do not develop such drug-related problems (Budney and Moore, 2002, Looby and Earleywine, 2007, Temple et al., 2011). This distinction is especially important since cannabis use per se is regarded to be relatively harmless (Nutt et al., 2010, Van Amsterdam et al., 2010). Furthermore, there are indications that the risk of comorbid disorders is substantially lower (often not exceeding the general population level) among non-dependent compared to dependent frequent cannabis users (Swift et al., 2001, Van der Pol et al., 2013b)
There were 73 incident cases of cannabis dependence within the three-year follow-up
one of the sociodemographic variables predicted incident cannabis dependence, except living alone. Baseline cannabis use variables that predicted incident cannabis dependence included a diagnosis of 12-month abuse, number of lifetime CUD symptoms, (also) using at daytime, continual smoking, and coping motives for cannabis consumption. Remarkably, cannabis exposure variables were not predictive of the transition from non-dependent to dependent cannabis use. Lifetime and current mental disorders were frequent in both family members and participants but these disorders were not predictive for the development of first incident cannabis dependence, and neither was family childhood adversity. However, the number of childhood adversities and the number of recent negative life events were associated with the development of dependence and so were two of the three impulsivity subscales. Finally, the lack of ‘heart unburdening’ social support was a significant predictor (p ≤ 0.10).
While dependent and non-dependent users were similar in this study, this comparison showed that both groups of frequent cannabis users differed from the general population particularly on risk factors previously reported in prospective general population studies. Together with the finding that the incidence of cannabis dependence in general population studies is strongly associated with frequent cannabis use, this may indicate that predictors from general population studies are mainly related to the transition from incidental to frequent cannabis use and not to the transition from frequent cannabis use to cannabis dependence
among frequent cannabis users cannabis consumption motives and habits are more important predictors of future dependence than the actual cannabis (THC) exposure level.
The number of negative life events was also an independent predictor; the risk of dependence increased with 43% with every additional negative event… A major financial problem was the strongest predictor among all negative life events
it should be noted that cultural differences may be important. For example, legal problems due to cannabis use are rare in the Netherlands (1.9% in the current sample) and no OR could be calculated, but legal problems might be an important predictor in countries with more restrictive regulations (Perkonigg et al., 2008)
Pope H Jr, Gruber A, Hudson J, Huestis M, Yurgelun-Todd D. 2001. “Neuropsychological performance in long-term cannabis users” Arch Gen Psychiatry 58(10): 909-15
http://www.ncbi.nlm.nih.gov/pubmed/11576028
At days 0, 1, and 7, current heavy users scored below controls on recall of word lists; by day 28, no differences among the groups on any test results, no associations between lifetime cannabis use and test scores.
Pope H Jr, Gruber A, Hudson J, Cohane G, Huestis M, Yurgelun-Todd D. 2003. “Early-onset cannabis use and cognitive deficits: what is the nature of the association?” Drug Alcohol Depend 69(3):303-10
http://www.ncbi.nlm.nih.gov/pubmed/12633916
Early-onset cannabis users exhibit poorer cognitive performance than late-onset users or control subjects, especially in verbal IQ when we adjusted for VIQ, virtually all differences between early-onset users and controls on test measures ceased to be significant.
Porter C, Lawson J, Bigler E. 2005. “Neurobehavioral sequelae of child sexual abuse.” Child Neuropsychol 11(2): 203-20
http://www.ncbi.nlm.nih.gov/pubmed/16036445
diminished attention/concentration performance
Power R, Verweij K, Zuhair M, Montgomery G, Henders A, Heath A, Madden P, Medland S, Wray N, Martin N. 2014. “Genetic predisposition to schizophrenia associated with increased use of cannabis” Molecular Psychiatry 19: 1201-1204
http://www.nature.com/mp/journal/v19/n11/full/mp201451a.html
In a sample of 2082 healthy individuals, we show an association between an individual’s burden of schizophrenia risk alleles and use of cannabis. This was significant both for comparing those who have ever versus never used cannabis (P=2.6 × 10−4), and for quantity of use within users (P=3.0 × 10−3)
part of the association between schizophrenia and cannabis is due to a shared genetic aetiology
Power B, Dragovic M, Badcock J, Morgan V, Castle D, Jablensky A, Stefanis N. 2015. “No additive effect of cannabis on cognition in schizophrenia” Schizophr Res 168(1-2): 245-51
http://www.ncbi.nlm.nih.gov/pubmed/26235754
Confounding variables underpin the association between cannabis use and cognitive function in psychotic disorders. Taken together, it would appear that cannabis use or dependence has no additive effect on cognitive dysfunction in these disorders
Price C, Hemmingsson T, Lewis G, Zammit S, Allebeck P. 2009. “Cannabis and suicide: longitudinal study” British Journal of Psychiatry 195: 492-497
http://bjp.rcpsych.org/content/195/6/492.full
Although there was a strong association between cannabis use and suicide, this was explained by markers of psychological and behavioural problems. These results suggest that cannabis use is unlikely to have a strong effect on risk of completed suicide, either directly or as a consequence of mental health problems secondary to its use
Proal AC, Fleming J, Galvez-Buccollini JA, DeLisi LE. 2014. “A controlled family study of cannabis users with and without psychosis” Schizophrenia Research 152.1 283-288
Cannabis using psychotics no more or less likely to have schizophrenic relatives than non-using psychotics; suggest that having an increased familial morbid risk for schizophrenia may be the underlying basis for schizophrenia in cannabis users and not cannabis use by itself.
we found a tendency for depression and bipolar disorder to be increased in the relatives of cannabis users in both the patient and control samples. This might suggest that cannabis users are more prone to affective disorders than their non-using samples or vice versa
we conclude that cannabis does not cause psychosis by itself. In genetically vulnerable individuals, while cannabis may modify the illness onset, severity and outcome, there is no evidence from this study that it can cause the psychosis
Quickfall J, Crockford D. 2006. “Brain neuroimaging in cannabis use: a review” J Neuropsychiatry Clin Neurosci 18(3): 318-32
http://neuro.psychiatryonline.org/article.aspx?articleid=102807
Rademacher D, Meier S, Shi L, Ho W, Jarrahian A, Hillard C. 2008. “Effects of acute and repeated restraint stress on endocannabinoid content in the amygdala, ventral striatum, and medial prefrontal cortex in mice” Neuropharmacology 54(1): 108-16
http://www.ncbi.nlm.nih.gov/pubmed/17675104/
Amygdalar N-arachidonylethanolamine was decreased after 1, 7, and 10 restraint episodes; 2-arachidonylglycerol was increased after the 10th restraint. A similar pattern occurred in the medial prefrontal cortex (mPFC): N-arachidonylethanolamine was decreased after the 7th and 10th restraints and 2-arachidonylglycerol was increased after the 10th restraint. In the ventral striatum, the pattern reversed: N-arachidonylethanolamine was increased after the 10th restraint and 2-arachidonylglycerol was decreased after the 7th restraint. Palmitoylethanolamide contents changed in parallel with N-arachidonylethanolamine in the amygdala and ventral striatum. A single restraint episode did not affect the activity of fatty acid amide hydrolase (FAAH) in any of the brain regions examined. After the 10th restraint, both V(max) and K(m) for N-arachidonylethanolamine were increased in the mPFC; while only the V(max) was increased in the amygdala. On the other hand, the V(max) of FAAH was decreased in ventral striatum after the 10th restraint. After the 10th restraint, the maximum velocity for 2-oleoylglycerol hydrolysis was increased in mPFC; no other changes in 2-oleoylglycerol hydrolysis occurred. Repeated exposure to restraint produced no changes in CB(1) receptor density in any of the areas examined. These studies are consistent with the hypothesis that stress exposure alters endocannabinoid signaling in the brain and that alterations in endocannabinoid signaling occur during habituation to stress
Ragland J, Laird A, Ranganath C, Blumenfeld R, Gonzales S, Glahn D. 2009. “Prefrontal activation deficits during episodic memory in schizophrenia” Am J Psychiatry 166(8): 863-874
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2885958/
Episodic memory impairments represent a core deficit in schizophrenia that severely limits patients’ functional outcome. This quantitative meta-analysis of functional imaging studies of episodic encoding and retrieval tests the prediction that these deficits are most consistently associated with dysfunction in the prefrontal cortex
18 whole-brain studies in standard space that included a healthy comparison sample and low-level baseline contrast were selected
patients showed less prefrontal activation than comparison subjects in the frontal pole, dorsolateral and ventrolateral prefrontal cortex during encoding, and the dorsolateral prefrontal cortex and ventrolateral prefrontal cortex during retrieval. The ventrolateral prefrontal cortex encoding deficits were not present in studies that provided patients with encoding strategies, but dorsolateral prefrontal cortex deficits remained and were not secondary to group performance differences. The only medial temporal lobe finding was relatively greater patient versus comparison subject activation in the parahippocampal gyrus during encoding and retrieval
there was no evidence of reduced hippocampal or surrounding medial temporal lobe activation in patients versus comparison subjects during encoding or retrieval. The only group difference in the medial temporal lobe was a relative increase in activation in patients in the parahippocampal gyrus during encoding and retrieval
Raison C, Capuron L, Miller A. 2006. “Cytokines sing the blues: inflammation and the pathogenesis of depression” Trends Immunol 27(1): 24-31
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3392963/
Stress, which can precipitate depression, can also promote inflammatory responses through effects on sympathetic and parasympathetic nervous system pathways. Finally, depression might be a behavioral byproduct of early adaptive advantages conferred by genes that promote inflammation
individual symptoms, especially sleep disturbance [40], have been reported to contribute disproportionately to the association between depression and inflammation
Hypersecretion of CRH is considered to be the crucial biological substrate of the well-known link between psychosocial stress and depression [42]
the well-characterized hyperactivity of the HPA axis and sympathetic nervous system in depression is believed to be secondary to CRH hyperactivity, in part because of impaired negative regulation of CRH by glucocorticoids [43]. Altered CRH regulation by glucocorticoids is thought to be a result of alterations in the functioning of the glucocorticoid receptor [44]. Changes in regional brain activity have also been documented in depression and involve abnormal metabolic activity in the prefrontal cortex and altered dopamine metabolism in the basal ganglia (caudate) [45,46]
Downstream cytokine signal transduction pathways, including mitogen-activated protein kinases (MAPKs) and nuclear factor κB (NF-κB), also disrupt glucocorticoid receptor signaling [55,56]
Induction of NF-κB in the brain following peripheral cytokine (IL-1) exposure has been shown to mediate many of the behavioral effects of IL-1, including social withdrawal and decreased food intake [58]. NF-κB induction in the brain also might contribute to alterations in neuronal growth and survival, especially through the induction of nitric oxide (e.g. via inducible nitric oxide synthase, iNOS) and, ultimately, oxidative stress, which has been shown to alter promoter function for several genes central to synaptic plasticity [59,60]
IFN-α is a potent inducer of proinflammatory cytokines, including IL-6 and, to lesser extent, IL-1 β and TNF-α [54]. Substantiating the fact that the IFN-α-induced behavioral syndrome is indeed depression as encountered in other venues (e.g. mental health clinics) is that IFN-α-induced depression: (i) is responsive to treatment with standard antidepressant therapy [61]; (ii) is associated with alterations in serotonin metabolism, in part related to increased activity of the metabolic enzyme indoleamine 2,3, dioxygenase, which degrades tryptophan to kynurenine, which is then metabolized to quinolinic acid [62]; and (iii) is associated with alterations in CRH function as manifested by an exaggerated adrenocorticotropic hormone (ACTH) and cortisol response to the first administration of IFN-α in patients who ultimately develop depression [63]
IFN-α has been shown to alter metabolic activity in the basal ganglia [64], possibly representing disruption of basal ganglia circuitry involving dopamine and subserving hedonic tone (i.e. the capacity to experience pleasure and reward), as well as psychomotor speed. IFN-α has also been found to alter fundamental information processing in the dorsal part of the anterior cingulate cortex (ACC), as revealed by functional magnetic resonance imaging. In comparison with control subjects, IFN-α-treated patients exhibited increased ACC activity in response to a task with a low error rate, possibly representing an increased sensitivity to negative events or conflict [65]
Inflammation is the sine qua non of pathology
In humans, acute stress (e.g. laboratory stressor) and chronic stress (including lack of social support) have both been associated with increased production and/or release of proinflammatory cytokines and decreases in anti-inflammatory cytokines such as IL-10 [70–72]
Rajesh M, Mukhopadhyay P, Batkai S, Hasko G, Liaudet L, Drel V, Obrosova I, Pacher P. 2007. “Cannabidiol attenuates high glucose-induced endothelial cell inflammatory response and barrier disruption” Am J Physiol Heart Circ Physiol 293(1): H610-H619
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2228254/
HG markedly increased mitochondrial superoxide generation (measured by flow cytometry using MitoSOX), NF-κB activation, nitrotyrosine formation, upregulation of iNOS and adhesion molecules ICAM-1 and VCAM-1, transendothelial migration of monocytes, and monocyte-endothelial adhesion in HCAECs. HG also decreased endothelial barrier function measured by increased permeability and diminished expression of vascular endothelial cadherin in HCAECs. Remarkably, all the above mentioned effects of HG were attenuated by CBD pretreatment
Rajesh M, Pan H, Mukhopadhyay P, Batkai S, Osei-Hyiaman D, Hasko G, Liaudet L, Gao B, Pacher P. 2007. “Pivotal advance: cannabinoid-2 receptor agonist HU-308 protects against hepatic ischemia/reperfusion injury by attenuating oxidative stress, inflammatory response, and apoptosis” J Leukoc Biol 82(6): 1382-1389
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2225476/?report=classic
The potent CB2 receptor agonist HU-308, given prior to the induction of I/R, significantly attenuated the extent of liver damage (measured by serum alanine aminotransferase and lactate dehydrogenase) and decreased serum and tissue TNF-α, MIP-1α, and MIP-2 levels, tissue lipid peroxidation, neutrophil infiltration, DNA fragmentation, and caspase 3 activity. The protective effect of HU-308 against liver damage was also preserved when given right after the ischemic episode. HU-308 also attenuated the TNF-α-induced ICAM-1 and VCAM-1 expression in HLSECs, which expressed CB2 receptors, and the adhesion of human neutrophils to HLSECs in vitro. These findings suggest that selective CB2 receptor agonists may represent a novel, protective strategy against I/R injury by attenuating oxidative stress, inflammatory response, and apoptosis
Ischemia reperfusion (I/R) is the major mechanism of organ injury during myocardial infarction, stroke, coronary bypass surgery, and organ transplantation. The devastating effects of I/R arise from the acute generation of reactive oxygen and nitrogen species following reoxygenation, which inflict direct tissue injury and initiate a chain of deleterious cellular responses leading to inflammation, cell death, and eventually, organ failure (reviewed in refs. [1–7])
Ranganathan M, Braley G, Pittman B, Cooper T, Perry E, Krystal J, D’Sousa D. 2009. “The effects of cannabinoids on serum cortisol and prolactin in humans” Psychopharmacology (Berl) 203(4): 737-744
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2863108/
40 frequent users and 36 healthy controls; subjects also met the criteria for current DSM-IV cannabis abuse disorder but not cannabis dependence… users were permitted to use cannabis until 24 h prior to each test day
Baseline cortisol levels were similar between the two groups; THC increased cortisol in both, but frequent users had a blunted response, driven by the highest dose
Frequent cannabis users had lower baseline prolactin levels; THC administration did not affect prolactin — variation entirely within normal physiological range
In rats, lower HPA axis activity is reported to be associated with heightened acquisition of drug self-administration (Kosten and Ambrosio 2002)
adolescents with early onset of cannabis use had lower basal cortisol levels (Huizink et al. 2006
speculation that prolactin may be decreased by enhancement of dopamine tone
Rajavashisth TB, Shaheen M, Norris KC, Pan D, Sinha SK, Ortega J, Friedman TC. 2012. “Decreased prevalance of diabetes in marijuana users: cross-sectional data from the National Health and Nutrition Examination Survey (NHANES) III” BMJ Open 2:e000494
Marijuana users had a lower age-adjusted prevalence of DM compared to non-marijuana users (OR 0.42, 95% CI 0.33 to 0.55; p<0.0001). The prevalence of elevated C reactive protein (>0.5 mg/dl) was significantly higher (p<0.0001) among non-marijuana users (18.9%) than among past (12.7%) or current light (15.8%) or heavy (9.2%) users. In a robust multivariate model controlling for socio-demographic factors, laboratory values and comorbidity, the lower odds of DM among marijuana users was significant (adjusted OR 0.36, 95% CI 0.24 to 0.55; p<0.0001)
Ramaekers J, Kauert G, Theunissen E, Toennes S, Moeller M. 2009. “Neurocognitive performance during acute THC intoxication in heavy and occasional cannabis users” J Psychopharmacol 23: 266
http://www.ncbi.nlm.nih.gov/pubmed/18719045
Twenty-four subjects (12 occasional cannabis users and 12 heavy cannabis users) participated in a double-blind, placebo-controlled, two-way mixed model design. Both groups received single doses of THC placebo and 500μ g/kg THC by smoking. Performance tests were conducted at regular intervals between 0 and 8 h after smoking, and included measures of perceptual motor control (critical tracking task), dual task processing (divided attention task), motor inhibition (stop signal task) and cognition (Tower of London). THC significantly impaired performance of occasional cannabis users on critical tracking, divided attention and the stop signal task. THC did not affect the performance of heavy cannabis users except in the stop signal task, i.e. stop reaction time increased, particularly at high THC concentrations. Group comparisons of overall performance in occasional and heavy users did not reveal any persistent performance differences due to residual THC in heavy users
Performance impairments are maximal during the first hour after smoking and sharply decline over 2 –4 h after THC use
Objective performance impairments have been shown to emerge at serum THC concentrations as low as 2 – 5 ng/ml, whereas maximal performance impairments have been observed at serum concentrations >30 ng/ml (Ramaekers, et al ., 2006c)
From a public health perspective, the major concern about the acute effects of cannabis is the possibility of accidents if users drive or operate machinery while intoxicated (Hall and Solowij, 1998)
tolerance developed more rapidly, while the subjects were constantly treated with the lower, more frequent oral doses (Jones, et al ., 1981). Thus, if the doses of THC are small and infrequent, little tolerance seems to develop… frequent or long-term users of cannabis may not develop substantial tolerance unless they are in a constant state of ‘ high ’ as evinced by steady-state THC concentrations in blood
frequent use of cannabis (>smoking on more than 4 days/week) during the previous year in case of heavy users (2.3 joints per occasion, 240 occasions per year) and weekly use or less in case of occasional cannabis users (1.2 joints per occasion, 55 occasions per year)… recruited through advertisements in coffee shops
Hart,et al. (2001), who reported the absence of any acute effects of smoking cigarettes containing 1.8% and 3.9% THC on complex cognitive func-tions in heavy, daily users of cannabis… all subjects tested positive to THC in urine prior to smoking the experimental cigarettes… may thus also have reflected a presence of cognitive impairment during both treatment conditions (Ramaekers et al., 2006a)
cigarettes were prepared beforehand for each individual from stock provided by the Dutch Bureau for Medicinal Cannabis. Marijuana cigarettes were prepared from batches containing 13% THC, a standard potency for marijuana sold at Dutch pharmacies for medical use… i.e. inhalation for 4 s, hold breath for 10 s and exhale/break for 15 s
puffs per cigarette: placebo and THC condition were 25 (6.4) and 28 (15.4), res-pectively, by occasional users and 22 (7.5) and 22 (8.2), respec-tively, by heavy users
Subjects were not allowed to use alcohol on the day prior to an experi-mental session and were requested to arrive at experimental sessions well rested… Subjects received a stan-dardized breakfast prior to smoking
The crit-ical tracking task (CTT) was conducted at 10 min, 3 h 05 min, 5 h 05 min and 7 h 05 min post-smoking; a divided attention task (DAT) was conducted at 20 min, 3 h 20 min, 5 h 20 min and 7 h 20 min post-dosing; the stop signal task (SST) was con-ducted at 35 min, 3 h 30 min, 5 h 30 min and 7 h 30 min post-smoking and a Tower of London (TOL) task was conducted at 60 min, 3 h 40 min, 5 h 40 min and 7 h 40 min post-dosing. Performance tasks were selected because of their demonstrated sensitivity to the impairing effects of THC (Ramaekers, et al ., 2006b)
The ‘ critical tracking test’ measures the subject’ s ability to con-trol a displayed error signal in a first-order compensatory tracking task. Error is displayed as a horizontal deviation of a cursor from the midpoint on a horizontal, linear scale. Com-pensatory joystick movements null the error by returning the cursor to the midpoint. The subject ’ s compensatory response increases in frequency with an increasing phase lag. Control is lost at the point where the compensatory response lags the cur-sor’ s last movement by 180°. The response frequency at this point is defined as the critical frequency or lambda-c. The test includes five trials of which the lowest and the highest scores were removed
THC increased Lambda-c in the CTT (F1,21 = 6,13; P = 0.022). However, THC induced impairment of critical tracking performance only occurred in occasional cannabis users when compared with heavy users as indicated by a signif-icant THC × cannabis use history interaction ( F 1,21 = 5,62; P = 0.027). In addition, cannabis use history also affected criti-cal tracking performance… worse in occasional cannabis users when compared with heavy users, independent of treatment condition ( F 1,21 = 5,28; P = 0.032)
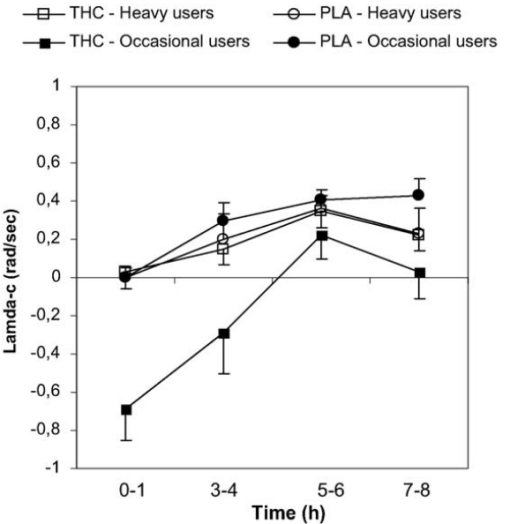
The ‘ divided attention task’ measures the subject’ s ability to divide attention between two tasks performed simultaneously. The primary task consists of tracking task as described above, but at a constant level of difficulty set at 50% of the subject’ s maximum capacity. Tracking error is measured as the differ-ence in mm between the position of the cursor and the midpoint of the scale
Performance in the DAT was significantly affected by THC. THC increased tracking error (F 1,21 = 7,38; P = 0.013) and the number of control losses ( F 1,21 = 5,28; P = 0.032) in the pri-mary task and decreased the number of hits in the secondary task ( F 1,21 = 5,93; P = 0.024). The overall effect of THC was most prominent during the first hour after smoking and only in occasional cannabis users relative to heavy users
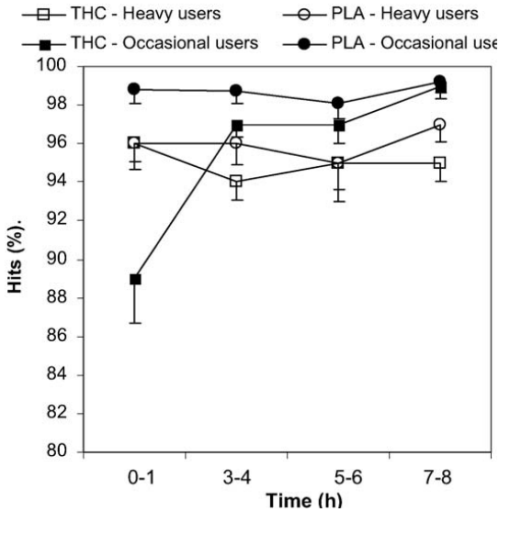
The ‘ stop signal task ’ measures motor impulsivity, which is defined as the inability to inhibit an activated or pre-cued response leading to errors of commission
In the SST, stop reaction time was affected by THC × time after smoking ( F 3,63 = 4,08; P = 0.01). Inspection of the data shows that stop reaction time primarily increased during the first hour of smoking. Accuracy of responses generally decreased following THC (F1,21 = 5,13; P = 0.034). Reaction time on go trials was not affected by THC
The ‘ Tower of London’ is a decision-making task that mea-sures executive function and planning (Shallice, 1982). The original version of the TOL consists of three coloured balls, which must be arranged on three sticks to match the target configuration on a picture, while only one ball can be moved at a time… The subject decides as quickly as possible, whether the end-arrangement can be accomplished in 2, 3, 4 or 5 steps
Performance in the TOL task was not affected by THC
THC concentrations were generally higher in heavy users than in occasional users during the THC condition. At baseline and throughout the placebo condition, heavy users displayed residual levels of THC ranging between 2 and 4 ng/ml (mean)… In heavy cannabis users, a single dose of 500 μ g/kg THC produced a mean peak concentration of 120.9 ng/ml, whereas the same dose produced a mean peak concentration of 49.1 ng/ml in occasional cannabis users
a single dose of 500 μg/kg THC impairs tracking performance, divided attention and inhibitory control in occasional cannabis users. Heavy cannabis users, however, did not display acute impairments on most of the performance tasks although impulse control decreased at high THC concentrations
direct comparisons of performance (independent of treatment condition) in occa-sional and heavy users revealed no significant differences in any of the performance tests, with the exception of the CTT… differences between tracking performance of both groups during placebo were always very small
Among occasional users, six out of 12 attested to ever driving under the influence of cannabis (DUIC). The average fre-quency of DUIC per year in occasional users, however, was relatively low, i.e. 0.5. Among heavy cannabis users, 11 of 12 attested to DUIC on a regular basis. Their mean frequency of DUIC was relatively high, i.e. about 94 times per year. Heavy cannabis users thus seem accustomed to driving under the influence of cannabis, whereas occasional users are not. Heavy users are likely to show up in the general driving popu-lation regularly as acute THC impairments are minor in this group. Occasional users, however, do not DUIC on a regular basis… crash risk calculations may be confounded by cannabis use history of drivers
no performance correlates were present for THC concentrations between 1 and 3 h after smoking when mean THC concentrations were between 6 and 19 ng/ml in heavy users and between 3 and 9 ng/ml in occasional users
Ramírez BG, Blázquez C, del Pulgar TG, Guzmán Manuel, de Ceballos ML. 2005. “Prevention of Alzheimer’s Disease Pathology by Cannabinoids: Neuroprotection Meditated by Blockade of Microglial Activation.” Journal of Neuroscience 25(8):1904-1913
Ranganathan M, D’Souza D. 2006. “The acute effects of cannabinoids on memory in humans: a review” Psychopharmacology 188:425-444
http://www.ncbi.nlm.nih.gov/pubmed/17019571
Acute administration of Δ -9-THC transiently impairs immedi ate a nd delayed free recal l o f infor mation presen ted after, but no t before , drug admi nistration in a dose- and delay- dependen t manne r. In parti cular, canna-binoid s increa se intrusi on error s. The se effects are more robust with the inha led a nd intravenous r ou te an d corres pond to peak drug levels
Δ -9-TH C has a long half- life of approximately 4 days (Johan sson et al. 1988). Its principal active metaboli te, 11-hydroxy -THC, is more potent than Δ -9-THC . The time course of 11-hydroxy-THC blood level s correlates well with the psychological effects of inhaled and oral Δ -9-TH C (reviewe d in Agur ell et al. 1986). Therefore, in relating cognitive or behavioral data with blood level s, both Δ -9-THC and 11-hydroxy-THC b lood leve ls need to be consi dered
Intravenous dosing follows the pharmacoki-netics and pharmacodynamics (Fig.1) of the inhaled route, though blood levels tend to be higher
Rapoport J, Addington A, Frangou S, Psych M. 2005. “The neurodevelopmental model of schizophrenia: update 2005” Mol Psychiatry 10(5): 434-49
http://www.nature.com/mp/journal/v10/n5/full/4001642a.html
With heritability around 80%, nongenetic factors impairing development must also be part of the model, and any model must also account for the wide range of age of onset
Rasic D, Weerasinghe S, Asbridge M, Langille D. 2013. “Longitudinal associations of cannabis and illicit drug use with depression, suicidal ideation and suicidal attempts among Nova Scotia high school students”
http://www.ncbi.nlm.nih.gov/pubmed/23041136 in dropbox
Nine hundred and seventy-six school students in four high schools in northern Nova Scotia – Illicit drug use, with and without accompanying cannabis use, among high school students increases the risk of depression, suicidal ideation and suicidal attempts. Heavy cannabis use alone predicts depression but not suicidal ideation or attempts. Association between marijuana and depression disappears upon controlling for confounds.
Rauch S, Whalen P, Shin L, McInerney S, Macklin M, Lasko N, Orr S, Pitman R. 2000. “Exaggerated amygdala response to masked facial stimuli in posttraumatic stress disorder: a functional MRI study” Biol Psychiatry 47(9): 769-76
http://www.ncbi.nlm.nih.gov/pubmed/10812035/
Read J, Perry B, Moskowitz A, Connolly J. 2001. “The Contribution of Early Traumatic Events to Schizophrenia in Some Patients: A Traumagenic Neurodevelopmental Model” Psychiatry 64(4): 319-345
http://childtrauma.org/wp-content/uploads/2013/09/contribution_early_traumatic_events_schizophrenia_perry.pdf in dropbox
In the last four decades, for every study on the relationship between child abuse or neglect and schizophrenia there have been 30 on the biochemistry of schizophrenia and 46 on the genetics
studies that retrospectively examine the childhoods of adults diagnosed schizophrenic tend to search only for evidence of behavioral dysfunction. Rather than researching what was occurring in the lives of these children, findings are explained in terms of the “constitutional vulnerability” underlying schizophrenia
The fact that the children diagnosed schizophrenic as adults were 2.7 times more likely than those who were not mentally ill as adults to have been in an institution or children’s home (a finding absent for those with affective psychosis as adults) did not lead to hypotheses about what might have been going on in these children’s lives to have caused these disruptions
17% diagnosed schizophrenic had suffered traumatic brain injury
HPA sensitization, impaired negative feedback
Compared to other psychiatric patients, those who suffered childhood physical abuse (CPA) or childhood sexual abuse (CSA) are more likely to attempt suicide, have earlier first admissions and longer and more frequent hospitalizations, spend more time in seclusion, receive more medication, and have higher global symptom severity (Beck and van der Kolk 1987; Beitchman et al. 1992; Briere, Woo, McRae, Foltz, and Sitzman 1997; Bryer, Nelson, Miller, and Krol 1987; Goff, Brot-man, Kindlon, Waites, and Amico 1991; Petti-grew and Burcham 1997; Read 1998; Read, Agar, Barker-Collo, Davies, and Moskowitz 2001; Sansonnet-Hayden, Haley, Marriage, and Fine 1987).
In 13 studies of “seriously mentally ill” women the percentage that experienced either CSA or CPA ranged from 45% to 92% (Goodman, Rosenberg, Mueser and Drake 1998)
73% of girls in a child and adolescent inpatient unit had suffered either CSA or CPA – Ito 1993
psychiatric inpatients tend to underreport abuse (Dill et al 1991; Read 1997)
Male inpatient CSA rates range from 22% to 39% (Jacobson and Herald 1990; Rose et al. 1991; Sansonnet-Hayden et al 1987; Wurr and Partridge 1996) and are at least double the rates of CSA in the general male population (Palmer et al 1994)
Mixed gender prevalence rates of child and adolescent inpatients: CSA 37%, CPA 44%, emotional abuse 52%, emotional neglect 31%, physical neglect 61%. CSA had mean age of onset of 8 years, duration 2.1 years, involved penetration or oral sex
Perceptual aberration scale scores, predictive of clinical psychoses, are 10 times more common in young adults who were maltreated as children (Berenbaum 1999)
4% of children raised by “healthy” adoptive families were diagnosed as “severe+psychotic,” compared to 34% in “disturbed” adoptive families — Tienari 1991 family dysfunction 7 times more explanatory than genetic predisposition
Among women inpatients diagnosed schizophrenic, 60% had suffered CSA (Friedman and Harrison 1984)
Read and Argyle (1999) found that the content of 54% of schizophrenic symptoms in adult inpatients who had been abused were obviously related to child abuse (ie, command hallucinations to self harm being the voice of the perpetrator)
After controlling for factors related to disruption and disadvantage in childhood, women whose CSA involved intercourse were 12 times more likely than nonabused females to have had psychiatric admissions (Mullen et al 1993)
Read J, Fink P, Rudegeair T, Felitti V, Whitfield C. 2008. “Child Maltreatment and Psychosis: A Return to a Genuinely Integrated Bio-Psycho-Social Model” Clinical Schizophrenia & Related Psychoses 2(3): 199-266
http://www.theannainstitute.org/ACE%20folder%20for%20website/55%20Child%20abuse%20and%20Psychosis.pdf in dropbox
the imbalance is worsening since the turn of the century, with research into biological causes increasing its share of research into schizophrenia, outweighing social causes by about fifteen to one (Table 1). Furthermore, of 1,284 publications about childhood schizophrenia only five (0.4%) relate to child abuse and eight (0.7%) to poverty
Patients who were abused as children are far more likely to try to kill themselves (11, 12, 14, 16). Even within samples of schizophrenia, those abused as children are more likely to attempt suicide (17, 18)
British children raised in economic deprivation are four times more likely to develop nonschizophrenic disorders; eight times more likely to become schizophrenic. Even without a family history of psychosis, deprived children still 7 times more likely (20)
A study of people with a first episode of psychosis found that they were 2.3 times more likely than a control group to have had their mothers die before they were sixteen
In Finland (27) and the United States (28), people whose pregnancies had been unwanted are 2.5 and 1.75 times more likely to be diagnosed with schizophrenia
in the year prior to hospitalization, 63% suffered violence by partners and 46% of those living at home were assaulted by family members (36). Sexual assaults as adults are experienced by the majority of female patients and about a quarter of male patients
(32, 37-41) Assaults also occur in mental health settings (42, 43)
sexual assault was significantly related to schizophrenia, but not to mania, depression, substance abuse or borderline personality disorder (44). In another study of female patients, physical assault was significantly related only to psychosis (45)
The weighted averages, accounting for variation in sample size, show that 64.5% of the women, and 55.5% of the men, had been subjected to either CSA or CPA. The combined rate is 60.2%
Estimating the prevalence of childhood maltreatment by using only CSA and CPA leads to gross underestimation A study of first episode schizophrenia-spectrum inpatients (57), which found CSA and CPA rates of 39% and 78%, respectively, also found
the following: childhood emotional abuse 94%, childhood emotional neglect 89%, and childhood physical neglect 89%. A study of eighty-seven inpatients with chronic psychosis found that all had suffered either neglect or emotional abuse as children (58)
A 2005 review (32) identified ten studies showing that people abused as children score higher than other people on the schizophrenia and paranoia scales of MMPI or SCL-90-R
78% of female outpatients diagnosed schizophrenic had CSA, compared to 30% of anxiety disorders, 42% of depressive disorder (25). Among child inpatients, 77% of those sexually abused were diagnosed psychotic, compared to 10% of the other children (60)
36% of those whose parents had both scored high on EE were now diagnosed with schizophrenia, compared to 0% if only one, or neither, parent was high EE (61) EE is a euphemism for parental hostility, criticism and intrusiveness
Some studies find that neglect and emotional abuse are even more strongly related to psychosis than CPA and CSA (58, 68, 69)
the strongest relationships with abuse and neglect appear to be for hallucinations, esp voices commenting, and paranoid delusions (32, 53, 69, 80). although hallucinations, delusions, and thought disorder were all predicted by child abuse, regression analysis revealed that only hallucinations were predicted by child abuse in the absence of abuse in adulthood (50). relationship between child abuse and hallucinations exists across diagnostic boundaries. A study of bipolar affective disorder found that those subjected to CSA were twice as likely to have auditory hallucinations in general and six times as likely to hear voices commenting (81)
sexual abuse (not all childhood) were 15.5 times more likely to be psychotic (91). Odds Ratios (ORs) for sexual abuse, and for the four other victimization events (see Table 5), were all higher than the ORs for the three other diagnostic groupings
child abuse were between 3.6 and 13 times more likely to be psychotic, depending on psychosis severity (92) 7.3 for most severe level of psychosis, after controlling for twelve variables
98, 99; OR for the adolescents who had experienced sexual trauma was 5.1, while the OR for those bullied once a week or more in the preceding year was 3.1. consistent with the U.K. study, which found an OR for bullying of 4.2 (91)
Childhood neglect was only predictive of visual hallucinations and, interestingly, CPA predicted only tactile hallucinations (101)
those who had been sexually molested were seven times more likely to experience delusions (102)
5.7 for sexual abuse and 2.1 for violence in the home (103) ORs of 2.5 for sexually molested and 4.2 for CPA
one or more psychotic symptoms were four times more likely to have suffered CSA, six times more likely to have suffered CPA and ten times more likely to have witnessed domestic violence (104)
eleven population-based studies, by seven independent research teams, using nine different samples drawn from six countries. All found higher levels of child maltreatment or neglect in their psychosis groups. Eight tested, all found, dose-response
within a schizophrenia sample, childhood trauma predicts who has abused substances (106)
2007 US: ORs for psychosis were 1.6 for one type of trauma only, 3.7 for three types of trauma and 30.2 for five types (103) 2007 UK: five or more types of trauma being 198 times more likely to be diagnosed psychotic than nonabused people
sexual trauma were 4.7 times more likely to have one type of psychotic experience, but 6.4 times more likely to experience two types (100)
source monitoring difficulty may serve the defensive function of protecting the person from the distress of realizing the true source, an inner memory of childhood trauma in the past, by experiencing it as an external event in the present (53)
within a sample of schizophrenia patients those abused as children, especially those emotionally abused, have greater HPA axis dysregulation, measured by cortisol levels, than their non-abused counterparts (130)
Reed P, Anthony J, Breslau N. 2007. “Incidence of drug problems in young adults exposed to trauma and posttraumatic stress disorder: do early life experiences and predispositions matter?” Arch Gen Psychiatry 64(12): 1435-42
http://www.ncbi.nlm.nih.gov/pubmed/18056552
Young adults (n = 988; aged 19-24 years) free of clinical features of DSM-IV drug use disorders at the first young adult assessment and therefore at risk for newly incident drug use disorders during the 1-year follow-up period
Prior PTSD (but not trauma only) was associated with excess risk for drug abuse or dependence (adjusted relative risk, 4.9; 95% confidence interval, 1.6-15.2) and emerging dependence problems (adjusted relative risk, 4.9; 95% confidence interval, 1.2-20.1) compared with the no-trauma group controlling for childhood factors. Subjects with PTSD also had a greater adjusted relative risk for drug abuse or dependence compared with subjects exposed to trauma only (adjusted relative risk, 2.0; 95% confidence interval, 1.1-3.8) controlling for childhood factors.
Reichenberg A, Caspi A, Harrington H, Houts R, Keefe R, Murray R, Poulton R, Moffitt T. 2010. “Static and Dynamic Cognitive Deficits in Childhood Preceding Adult Schizophrenia: A 30-Year Study”
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3552325/
Children who developed adult schizophrenia exhibited developmental deficits (i.e., static cognitive impairments that emerge early and remain stable) on tests indexing verbal and visual knowledge acquisition, reasoning, and conceptualization. In addition, these children exhibited developmental lags (i.e., growth that is slower relative to healthy comparison subjects) on tests indexing processing speed, attention, visual-spatial problem solving ability, and working memory
origins of schizophrenia include two interrelated developmental processes evident from childhood to early adolescence (ages 7–13 years). Children who will grow up to develop adult schizophrenia enter primary school struggling with verbal reasoning and lag further behind their peers in working memory, attention, and processing speed as they get older
individuals who develop adult schizophrenia exhibit an 8-point deficit (0.5 standard deviations) in their childhood IQ
Ribolsi M, Koch G, Magni V, Di Lorenzo G, Rubino I, Siracusano A, Centonze D. 2009. “Abnormal brain lateralization and connectivity in schizophrenia” Rev Neurosci 20(10: 61-70
http://www.ncbi.nlm.nih.gov/pubmed/19526734
Rieder C, Cicchetti D. 1989. “Organizational perspective on cognitive control functioning and cognitive-affective balance in maltreated children.”
http://psycnet.apa.org/index.cfm?fa=buy.optionToBuy&id=1989-29732-001
maltreated children showed poorer executive functioning even in emotionally neutral contexts. Executive functioning declined among maltreated children in contexts that primed for aggression.
Rieder S, Chauhan A, Singh U, Nagarkatti M, Nagarkatti P. 2010. “Cannabinoid-induced apoptosis in immune cells as a pathway to immunosuppression” Immunobiology 215(8): 598-605
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3005548/
activation of CB2 provides a novel therapeutic modality against inflammatory and autoimmune diseases as well as malignancies of the immune system
There is a ‘hierarchy’ of cannabinoid receptor expression within the immune system (B cells > Natural Killer cells > Monocytes > Neutrophils > CD8 leukocytes > CD4 leukocytes), and the level of expression is dependent on the stimulus and the activation state of the cell (Lee et al. 2001). For instance, stimulation of splenocytes with LPS led to CB2 mRNA downregulation, whereas CD40 co-stimulation resulted in CB2 mRNA upregulation (Lee et al. 2001).
Several studies have shown that cannabinoids negatively regulate phagocytosis, cell-spreading and antigen presentation by macrophages (Cabral and Mishkin 1989; Lopez-Cepero et al. 1986; McCoy et al. 1995)
cannabinoids exert their immunosupressive properties in four main ways: (1) induction of apoptosis, (2) inhibition of cell proliferation, (3) inhibition of cytokine and chemokine production, and (4) induction of regulatory T cells (T regs)
THC-induced apoptosis in splenocytes in a CB2 receptor-dependent way
Ajulemic acid (Aja) is a metabolite of THC, and it is a potent analgesic and anti-inflammatory agent (George et al. 2008). Aja belongs to a class of cannabinoids called carboxy tetrahydrocannabinols. In controlled trials of patients with neuropathic pain, Aja proved as effective as morphine and in rats, it significantly reduced severity of adjuvant-induced arthritis (Karst et al. 2003; Zurier et al. 1998). One study investigated the effects of Aja on human peripheral blood T lymphocytes. This study showed that Aja reduced T cell proliferation by ∼75%, and induced apoptosis in a dose-dependent manner (at 1, 3 and 10 μM)
In the blood of patients with septic shock, 4-fold increase in AEA and 2-AG was detected (Wang et al. 2001), and this was also seen in LPS-induced hypotension (Varga et al. 1998)
Overall, there exists a biphasic role for cannabinoids, and it depends on three key factors: (1) the type of cannabinoid, (2) the dose of cannabinoid, and (3) the type of cell that the cannabinoid is acting on. The degree of cannabinoid receptor expression may play role in some cases; however, the receptor-independent mechanism of action is evident for cannabinoids in some cell populations
Riesco Y, Pérez Urdániz A, Rubio V, Izquierdo J, Sánchez Iglesias S, Santos J, Carrasco J. 1998. “The evaluation of personality disorders among inmates by IPDE and MMPI” Actas Luso Esp Neurol Psiquiatr Cienc Afines 26(3): 151-4
http://www.ncbi.nlm.nih.gov/pubmed/9717339/
A sample of 56 prisoners from a Spanish prison was studied, mean age 22, all male, 98% of them had never requested psychiatric help. The crimes most frequently committed were related to drug traffic and drug abuse (thefts, robberies, crimes against public health). There were also cases of homicide, homicide attempt, rape and kidnapping. 91% of the studied sample presented one or more personality disorders, being the most frequent: Antisocial (79%), Paranoid (52%) and Borderline (41%). The MMPI scales most frequently obtained were: Psychopathic deviation (59%), Paranoia (46%) and Schizophrenia (41%). There was a good clinical correlation between the IPDE and the MMPI results.
Ripoll N, Bronnec M, Bourin M. 2004. “Nicotinic receptors and schizophrenia” Curr Med Res Opin 20(7): 1057-74
http://www.ncbi.nlm.nih.gov/pubmed/15265251
smoking is very high in non-schizophrenic subjects presenting various psychiatric disorders (35 to 54%). However, the incidence of smoking is extremely high in schizophrenic patients: 80% to 90%, versus 25% to 30% of the general population
tobacco transiently restores the schizophrenic patient’s cognitive and sensory deficits. Smoking cessation also appears to exacerbate the symptoms of the disease Post-mortem binding studies have revealed a disturbance of nicotinic receptor expression, affecting the alpha(7) and alpha(4)beta(2) subunits, in various cerebral areas. Genetic linkage studies have also shown that the alpha(7) subunit is involved in schizophrenia. This review assesses the involvement of the nicotinic system in schizophrenia and suggests ways in which this system may participate in the pathophysiology of this disease.
Rivera P, Blanco E, Bindila L, Alen F, Vargas A, Rubio L, Pavon F, Serrano A, Lutz B, Rodriguez de Fonseca F, Suarez J. 2015. “Pharmacological activation of CB2 receptors counteracts the deleterious effect of ethanol on cell proliferation in the main neurogenic zones of the adult rat brain.” Front Cell Neurosci 9: 379
http://www.ncbi.nlm.nih.gov/pubmed/26483633
These results indicated that the specific activation of CB2 receptors rescued alcohol-induced impaired NPC proliferation, which is a potential clinical interest for the risk of neural damage in alcohol dependence.
Roberto M, Cruz M, Bajo M, Siggins G, Parsons L, Schweitzer P. 2010. “The endocannabinoid system tonically regulates inhibitory transmission and depresses the effect of ethanol in central amygdala” Neuropsychopharmacology 35(9): 1962-1972
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2904853/
Superfusion of the cannabinoid receptor (CB1) agonist WIN55212-2 (WIN2) onto central amygdala (CeA) neurons decreased evoked GABAA receptor-mediated inhibitory postsynaptic potentials (IPSPs) in a concentration-dependent manner, an effect prevented by the CB1 antagonists Rimonabant (SR141716, SR1) and AM251. SR1 or AM251 applied alone augmented IPSPs, revealing a tonic eCB activity that decreased inhibitory transmission in CeA. Paired-pulse analysis suggested a presynaptic CB1 mechanism. Intracellular BAPTA abolished the ability of AM251 to augment IPSPs, demonstrating the eCB-driven nature and postsynaptic origin of the tonic CB1-dependent control of GABA release. Superfusion of ethanol increased IPSPs and addition of WIN2 reversed the ethanol effect. Similarly, previous superfusion of WIN2 prevented subsequent ethanol effects on GABAergic transmission. The ethanol-induced augmentation of IPSPs was additive to CB1 blockade, ruling out a participation of CB1 in the action of acute ethanol. Our study points to an important role of CB1 in CeA in which the eCBs tonically regulate neuronal activity, and suggests a potent mechanism for modulating CeA tone during challenge with ethanol
In CeA slices, ethanol increases GABAergic transmission by acting principally at a presynaptic site to augment GABA release (Roberto et al, 2003; Nie et al, 2004)
Administration of a CB1 antagonist reduces alcohol self-administration and preference (Colombo et al, 2005), and mice deficient in CB1 consume less ethanol than do wild-type mice (Wang et al, 2003; Hungund et al, 2003). Conversely, CB1 activation increases ethanol consumption (Colombo et al, 2002)
apparent low levels of CB1 appear to be sufficient to generate a large physiological response. A similar profile has been observed in the basolateral amygdala and hippocampus where low levels of CB1 immunoreactivity were detected on glutamatergic neurons (and sometimes no immunoreactivity at all), yet robust and widespread CB1 physiological effects were observed in these neurons. Such results underlie the uncertainty of correlating the strength of CB1 staining with that of the physiological response
The implication of other nuclei is also unlikely because most connections between amygdaloid nuclei are glutamatergic (Sah et al, 2003), and this study involves GABAergic responses
CeA neurons release eCBs in response to elevated calcium, and the tonic inhibition of GABA release is homosynaptically regulated by the postsynaptic neuron
The fine-tuning of the GABAergic inhibitory system in CeA is a prerequisite for controlling the output neurons, and an active CB1 tone will dampen local GABA release and consequently increase the activity of CeA neurons… tonic CB1 activity could increase feed-forward inhibition to downstream nuclei. Conversely, administration of a CB1 antagonist in CeA will increase local inhibition and mute the CeA, consequently relieving inhibition of (disinhibiting) downstream structures
we predict that CB1 activation… increases the outflow of information to downstream structures
WIN2 is among the most potent CB1 agonists and its effects are much more pronounced than those of AEA or 2-AG on synaptic activity (Bajo et al, 2009). Thus, the physiological alteration of alcohol effects by eCBs in CeA is likely to be modulatory instead of preventive, and the effects of ethanol could be amplified or lessened according to ambient eCB levels at the time of alcohol consumption
SR1 reverses the ethanol-induced depression of excitatory transmission in cultured hippocampal neurons (Basavarajappa et al, 2008)
ethanol increased GABAergic transmission in neurons loaded with BAPTA to suppress the effect of CB1 blockade, further confirming that ethanol actions do not implicate the eCBs in CeA
CB1 and ethanol may act on common signaling pathways to regulate the release of GABA, as suggested in the cerebellum where activation of presynaptic protein kinase A (PKA) is necessary for the ethanol-induced increase in spontaneous GABA release (Kelm et al, 2008). CB1 activation is well known to inhibit adenylate cyclase and decrease PKA activity, and the mechanism of the cannabinoid-induced decrease of spontaneous GABA release in cerebellar neurons seems to involve PKA (Kelm et al, 2008). Therefore, the adenylate cyclase/PKA pathway may be a common signal transduction pathway targeted in opposite ways by eCBs and ethanol to regulate GABA release. Another potential candidate is protein kinase C epsilon (PKCɛ). We recently determined that a PKCɛ signaling pathway is activated by ethanol to augment GABA release in CeA (Bajo et al, 2008). PKCɛ reportedly negatively regulates the affinity of WIN2 at CB1 (Wallace et al, 2009), although a possible modulation of eCB activity at CB1 by PKCɛ is uncertain. Nonetheless, these two kinases seem to be interesting candidates to participate in the opposite regulation of GABA release by ethanol and eCBs, and a combined crosstalk between PKA and PKCɛ to regulate ethanol and eCB effects is also possible (Kelm et al, 2008)
Robinson D, Woerner M, McMeniman M, Mendolowitz A, Bilder R. 2004. “Symptomatic and functional recovery from a first episode of schizophrenia or schizoaffective disorder” Am J Psychiatry 161(3): 473-9
http://www.ncbi.nlm.nih.gov/pubmed/14992973
After 5 years, 47.2% (95% CI=36.0%-58.4%) of the subjects achieved symptom remission, and 25.5% (95% CI=16.1%-34.7%) had adequate social functioning for 2 years or more. Only 13.7% (95% CI=6.4%-20.9%) of subjects met full recovery criteria for 2 years or longer. Better cognitive functioning at stabilization was associated with full recovery, adequate social/vocational functioning, and symptom remission. Shorter duration of psychosis before study entry predicted both full recovery and symptom remission. More cerebral asymmetry was associated with full recovery and adequate social/vocational functioning; a schizoaffective diagnosis predicted symptom remission
Rodondi N, Pletcher MJ, Liu K, Hulley SB, Sidney S. 2006. “Marijuana use, diet, body mass index and cardiovascular risk factors (from the CARDIA study)” American Journal of Cardiology 15;98(4): 478-84
Rodríguez-Arias M, Navarrete F, Blanco-Gandia M, Arenas M, Aguilar M, Bartoll-Andres A, Valverde O, Minarro J, Manzanares J. 2015. “Role of CB2 receptors in social and aggressive behavior in male mice” Psychopharmacology (Berl) 232(16): 2019-31
http://www.ncbi.nlm.nih.gov/pubmed/25921034
Group-housed CB2KO mice exhibited higher levels of aggression in the social interaction test and displayed more aggression than resident WT mice. Isolation increased aggressive behavior in WT mice but did not affect CB2KO animals; however, the latter mice exhibited higher levels of social interaction with their WT counterparts… CB2 agonist JWH133 significantly reduced the level of aggression in aggressive isolated OF1 mice, an effect that decreased after pretreatment with the CB2 receptor antagonist AM630
Rohof W, Aronica E, Beaumont H, Troost D, Boeckxstaens G. 2012. “Localization of mGluR5, GABAB, GABAA, and cannabinoid receptors on the vago-vagal reflex pathway responsible for transient lower esophageal sphincter relaxation in humans: an immunohistochemical study” Neurogastroenterol Motil 24(4): 383-e173
http://www.ncbi.nlm.nih.gov/pubmed/22256945
Human mGluR5, GABA(A,B), and CB(1,2) receptors are abundantly expressed along the vago-vagal neural pathway and involved in the triggering of TLESRs
Romero J, Garcia L, Fernandez-Ruiz J, Cebeira M, Ramos J. 1995. “Changes in rat brain cannabinoid binding sites after acute or chronic exposure to their endogenous agonist, anandamide, or to delta 9-tetrahydrocannabinol” Pharmacol Biochem Behav 51(4): 731-7
http://www.ncbi.nlm.nih.gov/pubmed/7675852
upregulation
Rosen J, Schulkin J. 1998. “From normal fear to pathological anxiety” Psychol Rev 105(2): 325-50
http://www.ncbi.nlm.nih.gov/pubmed/9577241/
Pathological anxiety is conceptualized as an exaggerated fear state in which hyperexcitability of fear circuits that include the amygdala and extended amygdala (i.e., bed nucleus of the stria terminalis) is expressed as hypervigilance and increased behavioral responsivity to fearful stimuli. Reduced thresholds for activation and hyperexcitability in fear circuits develop through sensitization- or kindling-like processes that involve neuropeptides, hormones, and other proteins. Hyperexcitability in fear circuits is expressed as pathological anxiety that is manifested in the various anxiety disorders.
Rossi S, de Chiara V, Musella A, Kusayanagi H, Mataluni G, Bernardi G, Usiello A, Centonze D. 2008. “Chronic psychoemotional stress impairs cannabinoid-receptor-mediated control of GABA transmission in the striatum” J Neurosci 28(29): 7284-92
http://www.ncbi.nlm.nih.gov/pubmed/18632932/
inhibition of GABAergic IPSCs induced by the cannabinoid CB(1) receptor agonist HU210… was reduced after a single stressful episode and fully abolished after 3 and 7 d of stress exposure. Repeated psychoemotional stress also impaired the sensitivity of GABA synapses to endocannabinoids mobilized by group I metabotropic glutamate receptor stimulation, whereas the cannabinoid CB(1)-mediated control of glutamate transmission was unaffected by repeated exposure to an aggressor. Corticosteroids released in response to the activation of the hypothalamic-pituitary-adrenal axis played a major role in the synaptic defects observed in stressed animals, because these alterations were fully prevented by pharmacological blockade of glucocorticoid receptors and were mimicked by corticosterone injections. The recovery of stress-induced synaptic defects was favored when stressed mice were given access to a running wheel or to sucrose consumption, which function as potent natural rewards. A similar rescuing effect was obtained by a single injection of cocaine, a psychostimulant with strong rewarding properties. Targeting cannabinoid CB(1) receptors or endocannabinoid metabolism might be a valuable option to treat stress-associated neuropsychiatric conditions
Roulette C, Kazanji M, Breurec S, Hagen E. 2015. “High prevalence of cannabis use among Aka foragers of the Congo Basin and its possible relationship to helminthiasis” American Journal of Human Biology DOI: 10.1002/ajhb.22740
The prevalence of cannabis use among adult Aka men was high when compared to most global populations. THCA levels were negatively correlated with parasite infection and reinfection, supporting the self-medication hypothesis
Rubino T, Realini N, Castiglioni C, Guidali C, Vigano D, Marras E, Petrosino S, Perletti G, Maccarrone M, Di Marzo V, Parolaro D. 2008. “Role in anxiety behavior of the endocannabinoid system in the prefrontal cortex” Cereb Cortex 18(6): 1292-1301
http://cercor.oxfordjournals.org/content/18/6/1292.long
Low doses of the metabolically stable AEA analog, methanandamide, microinjected into the PFC, produced an anxiolytic-like response in rats, whereas higher doses induced anxiety-like behaviors
anxiolytic effect evoked by AEA might be due to the interaction with the CB1 cannabinoid receptor, whereas vanilloid receptors seem to be involved in AEA anxiogenic action
more marked increases or decreases of this endocannabinoid might lead to an anxiogenic response due to TRPV1 stimulation or the lack of CB1 activation, respectively
Increased neural activity was found in the orbitofrontal cortex and the anterior and posterior cingulate in subjects suffering from panic disorder (Bystritsky et al. 2001) and specific phobias (Dilger et al. 2003). Decreased binding of inhibitory γ-aminobutyric acid A (GABAA) receptors in the PFC was detected in panic and posttraumatic stress disorders (Malizia et al. 1998; Bremner et al. 2000)
we decided to microinject 2-AG in the PFC to assess the activity of this compound on anxiety behavior. As reported inFigure 8, at all tested doses (0.1–10 μg) 2-AG did not significantly modify the anxiety response in rats.
tonic activation of CB1 and, at higher concentrations, TRPV1 receptors by AEA, but not 2-AG, control anxiety in opposite ways. If, for any reason, the tissue levels of AEA become either too low or too high, thus leading to lack of CB1 activation or TRPV1 stimulation, respectively, anxiogenic responses are observed
Ruehle S, Rey A, Remmers F, Lutz B. 2012. “The endocannabinoid system in anxiety, fear memory and habituation” J Psychopharmacol 26(1): 23-39
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3267552/
a bidirectional influence of the ECS has been reported, whereby anxiolytic and anxiogenic responses have been obtained after both increases and decreases of the endocannabinoid tone
In fear memory paradigms, the ECS is mostly involved in the two opposing processes of reconsolidation and extinction of the fear memory. Whereas ECS activation deteriorates reconsolidation, proper extinction depends on intact CB1 receptor signalling. Thus, both for anxiety and fear memory processing, endocannabinoid signalling may ensure an appropriate reaction to stressful events. Therefore, the ECS can be considered as a regulatory buffer system for emotional responses
CB1 receptor agonists are reported to induce biphasic effects, with lower doses being anxiolytic and higher doses being anxiogenic (Viveros et al., 2005)
the same dose of delta 9-tetrahydrocannabinol (Δ9-THC) is able to promote anxiolytic responses when injected in the prefrontal cortex (PFC), whereas microinjections in the basolateral amygdala (BLA) lead to an anxiogenic response
contradictory anxiety-like results obtained in rats and mice can be explained by the different cannabinoid responsiveness of GABAergic and glutamatergic neurons in these two species
significant differences between CB1 receptor knockout animals and their wildtype littermates can only be found under aversive conditions (Haller et al., 2004a; Jacob et al., 2009). Therefore, the light conditions during the test and the housing and handling prior to the test are important parameters to be kept well-defined in experiments involving anxiety and the ECS
a different basal activation has been demonstrated for CB1 receptors expressed on glutamatergic neurons and GABAergic neurons, being higher on the latter
the great majority of the CB1 receptor are located on GABAergic neurons in the brain
CB1 receptor antagonists such as AM251 and SR141716 have been shown to increase inhibitory transmission in the central nucleus of the amygdala (CeA), suggesting a tonic activation of CB1 receptors on GABAergic neurons under basal conditions (Roberto et al., 2010)
increases of glutamate release induced by AM251 were never higher than 110% of the tonic release (Slanina and Schweitzer, 2005), whereas augmentation of GABAergic release after treatment with the same CB1 receptor antagonist reached levels of 130–140% (Roberto et al., 2010). Hence, the tonic inhibition mediated by the CB1 receptor is much more relevant on GABAergic synapses than on glutamatergic synapses
excitatory transmission has been estimated to be around 30-fold less sensitive to cannabinoids than inhibitory transmission (Ohno-Shosaku et al., 2002)
The group (of inhibitory synapses) with higher sensitivity is formed by perisomatically projecting basket cells (BC), whereas the dendritically projecting Schaffer-collateral associated cells (SCA) belong to the group with lower sensitivity to WIN55,212-2 (Lee et al., 2010)
A recent study revealed that the capacity to recruit G proteins is more prominent in CB1 receptors located on glutamatergic terminals than on GABAergic terminals (Steindel et al., 2008), suggesting a compensatory effect to the lower abundance of CB1 receptors on glutamatergic terminals
GABAergic localization, together with high basal activation and high sensitivity would confer an important role in the development of anxiogenic responses to cannabinoids to the CB1 receptor. On the other hand, glutamatergic localization, low basal activation and low sensitivity are factors that give the CB1 receptor an essential function in the development of anxiolytic responses to cannabinoids
rats – Local (Page et al., 2008) and systemic (Page et al., 2007) administration of the CB1 receptor agonist WIN55,212-2 causes an increase in extracellular norepinephrine (NE) in the PFC- the increase in NE release was accompanied by enhanced expression of tyrosine hydroxylase, the rate-limiting enzyme in the biosynthesis of catecholamine neurotransmitters (Boundy et al., 1998)
antagonists of the serotonergic type 1A (5-HT1A; WAY100635) and 5-HT2A/2C (ketanserin) receptors attenuated the CB1 receptor-dependent increase of corticosterone secretion. Thus, the anxiogenic action of the ECS through monoaminergic transmission appears to be related not only to NE but also to 5-HT
it can be hypothesized that cannabinoid enhancement of NE transmission is commonly associated with anxiogenic responses, whereas cannabinoid-mediated stimulation of serotonergic signalling is associated with anxiolytic responses after mild activation (FAAH inhibition, low doses of WIN55,212-2) and to anxiogenic-like reactions after stronger activation (HU-210) (McLaughlin et al., 2009)
The presence of cannabinoid type 2 (CB2) receptor in brain tissue has recently been related to feeding behaviour (Ishiguro et al., 2010), pain processing (Bingham et al., 2007), drug abuse and depression (Onaivi et al., 2008)
2-AG-mediated suppression of inhibitory transmission was not blocked by the CB1 receptor antagonist LY320135, but with the CB2 receptor antagonist AM630 (Morgan et al., 2009)
TRPV1 was required for the anxiogenic effect of high doses of the CB1 receptor agonist WIN55,212-2 (Campos and Guimaraes, 2009)
The main effect of SSRIs is achieved by the enhancement of serotonin activity, which involves the desensitization of 5-HT1A inhibitory autoreceptors (Nutt and Stein, 2006)
5-HT1Areceptors are possibly involved in the anxiolytic effects of CBD, as demonstrated by microinjections of low doses of CBD in the PAG area, which caused anxiolytic-like effects that are counteracted by the 5-HT1Areceptor antagonist WAY-100635, but not by the CB1 receptor antagonist AM251 (Campos and Guimaraes, 2008) (Russo et al., 2005)
CCK2 agonists are anxiogenic, and CCK2 antagonists reduce potentiated states of anxiety but do not appear to affect baseline anxiety responses (Rotzinger and Vaccarino, 2003)
stress alters endocannabinoid content in limbic areas and PFC (Rademacher et al., 2008). Further experiments have confirmed that chronic psychoemotional stress (viz. social defeat) blocks the normal reduction of inhibitory postsynaptic potentials (IPSPs) produced after application of the CB1 receptor agonist HU-210 to corticostriatal slices of C57BL/6 mouse brains (Rossi et al., 2008)… counteracted by pretreatment with CB1 and glucocorticoid receptor antagonists, revealing the importance of HPA-axis reactivity in this process
stress did not affect the normal CB1 receptor-mediated reduction of excitatory postsynaptic potentials (EPSPs), suggesting a more static profile of the CB1 receptor on glutamatergic terminals
increase of AEA levels by genetic or pharmacological inactivation of its degrading enzyme was able to reverse the stress-induced effect, and this response was also mediated by the CB1 receptor (Rossi et al., 2010). … Campos et al. (2010) concluded that an anxiogenic increase of AEA turned out to be anxiolytic (tested on the elevated plus-maze), when the animals were pre-exposed to restraint stress. Consequently, it can be hypothesized that the anxiogenic properties of the enhancement of AEA signalling (via AM404) rely on the activation of CB1 receptors on GABAergic terminals
stress-mediated downregulation of these CB1 receptors results in a stronger activation of CB1 receptors on glutamatergic terminals, leading to an anxiolytic response
long-lasting CB1 receptor downregulation on GABAergic terminals leads to a persistent increase in the strength of GABAergic inhibition of the glutamatergic transmission. This model represents a precise buffer system for the homeostasis of emotions
stimuli known to be activators of the reward system, such as voluntary exercise or sucrose consumption in rodents, were responsible for a larger reduction of the IPSPs by the CB1 receptor agonist HU-210 (De Chiara et al., 2010). Notably, this increase in CB1 receptor-dependent inhibition of IPSPs was not shown on EPSPs, and it was prevented by previous exposure to psychoemotional stress
chronic cocaine administration was shown to induce sensitization of GABAergic synapses (unlike glutamatergic) to the stimulation of the CB1 receptor (Centonze et al., 2007)
possible involvement of the ECS in early phases of fear memory processing was also suggested by the finding of a 20% increase in CB1 receptor mRNA expression in the IL cortex 48 hours after contextual fear conditioning (Lisboa et al., 2010)
reduced ECS activity during acquisition and retrieval seems to increase the subsequent fear response, whereas reduced ECS activity during consolidation may reduce the subsequent fear response.
Post-training (consolidation) and pre-test (retrieval) manipulations seem to have opposite effects, with the former increasing fear response when in the BLA or hippocampus and reducing fear upon systemic injections; whereas ECS activation during retrieval seems to reduce fear when in the BLA, hippocampus, IL or PAG and to enhance fear expression when systemic or in the PL
hypo-activation of the ECS by hippocampal AM251 injections improved reconsolidation (de Oliveira Alvares et al., 2008a), and hyperactivation by AEA, WIN55,212-2 or HU-210 injections in rats all reduced reconsolidation of the fear memory. These effects were independent of injection site (hippocampus, amygdala or insular cortex), paradigm (cued or contextual fear conditioning, or conditioned taste aversion) and time of measurement (at re-exposure, after re-instatement or spontaneous recovery after 7 days) (Lin et al., 2006; Kobilo et al., 2007; de Oliveira Alvares et al., 2008a). These studies all point to an amnesic role of endocannabinoid signalling. Therefore, it seems likely that activation of the ECS reduces the reconsolidation of fear memories, whereas hypo-activation of the ECS promotes their reconsolidation, and thereby leads to enduring fear responses
endocannabinoid signalling is dispensable for extinction of appetitive memories (e.g. Niyuhire et al., 2007; Manwell et al., 2009), the role of the endocannabinoids in extinction seems to be specific for aversive memories.
the fear-reducing effect of the ECS highly depends on the strength of the harmful stimulus encountered (Kamprath et al., 2009)… The fear-alleviating effect of endocannabinoids depended on endocannabinoid-driven modulation of glutamatergic transmission
stress habituation mediated by the ECS also crucially involves the modulation of glutamatergic neurotransmission (Patel and Hillard, 2008)
the ECS is crucial for efficient extinction learning. Therefore, it is tempting to hypothesize that the activation of the CB1 receptor would improve extinction, but the available data reveal a more complicated picture
injection of AEA into the hippocampus of rats directly after an extinction session led to improved extinction retention on the following day (de Oliveira Alvares et al., 2008a)
Chronic treatment with a CB1 receptor agonist leads to impaired extinction as shown in a study by Ashton et al. (2008), demonstrating that treatment with a high dose of Δ9-THC over 6 days strongly delayed cued extinction
treatment with CB1 receptor agonist improves extinction in rats when administered in a low dose, whereas it has no effect or even delays extinction when a high dose or chronic treatment is used. This effect of high dose or chronic agonist treatment might be due to internalization/desensitization of the CB1 receptor upon strong or continuous activation (Wu et al., 2008), leading to a resistance to exogenous as well as endogenous cannabinoids and therefore having the same effect as blocking CB1 receptor signalling
Katona and Freund (2008) GABAergic CB1 receptors as members of a regular buffer of inhibitory transmission, in opposition to the glutamatergic CB1 receptors, which have the task to reduce excitatory transmission only after excessive glutamate release
Considering the predominant action of stress/reward-mediated plasticity on GABAergic transmission, the CB1 receptors on GABAergic terminals could be considered as a dynamic element that can be altered in order to continuously maintain an appropriate and optimal emotional reactivity. The CB1 receptors on glutamatergic terminals, on the other hand, would be required only in situations of more prominent GABA-glutamate unbalance
increases of NE release in the locus coeruleus and PFC commonly led to anxiogenic responses, whereas serotonergic enhancement in the dorsal raphe nucleus and PFC induced anxiolytic responses accompanied by increased sociability and antidepressant-like behaviours
it may be postulated that the ECS tone determines the balance between the processes of maintaining or strengthening the original memory (reconsolidation) and the establishment of a new memory or habituation to the situation (extinction). In this way, the ECS may protect the organism from over-reaction to aversive events. This suggests that endocannabinoid-mediated plasticity may play a similar role in fear memory-processing as in anxiety, namely to ensure an appropriate reaction to negative events. Thus, the ECS may be considered as a regulatory buffer system for emotional responses.
Ruiz-Veguilla M, Gurpegui M, Barrigon M, Ferrin M, Marin E, Rubio J, Gutierrez B, Pintor A, Cervilla J. 2009. “Fewer neurological soft signs among first episode psychosis patients with heavy cannabis use” Schizophr Res 107(2-3): 158-64
http://www.ncbi.nlm.nih.gov/pubmed/18805673
The presence of high NSS showed a significant independent association with not having been a heavy cannabis user (OR=8.3; 95% CI, 2.4-33.3), family history of psychosis (OR=4.3; 95% CI, 1.2-14.9), male sex (OR=4.0; 95% CI, 1.2-14.0), lower score in verbal fluency and higher score in negative symptoms (both p<0.01).
Russo, E. 2011. “Taming THC: potential cannabis synergy and phtocannabinoid-terpenoid entourage effects” British Journal of Pharmacology 163: 1344-1364
http://www.ncbi.nlm.nih.gov/pubmed/21749363 in dropbox
Sabia J, Swigert J, Young T. 2015. “The effect of medical marijuana laws on body weight” Health Economics DOI: 10.1002/hec.3267
http://onlinelibrary.wiley.com/doi/10.1002/hec.3267/abstract
enforcement of MMLs is associated with a 2% to 6% decline in the probability of obesity. We find some evidence of age-specific heterogeneity in mechanisms. For older individuals, MML-induced increases in physical mobility may be a relatively important channel, while for younger individuals, a reduction in consumption of alcohol, a substitute for marijuana, appears more important. These findings are consistent with the hypothesis that MMLs may be more likely to induce marijuana use for health-related reasons among older individuals, and cause substitution toward lower-calorie recreational ‘highs’ among younger individuals. Our estimates suggest that MMLs induce a $58 to $115 per-person annual reduction in obesity-related medical costs.
Sagar K, Dahlgren M, Gonenc A, Racine M, Dreman M, Gruber S. 2015. “The impact of initiation: early onset marijuana smokers demonstrate altered Stroop performance and brain activation” Developmental Cognitive Neuroscience in press
http://www.sciencedirect.com/science/article/pii/S1878929315000407
50 chronic MJ smokers divided into early onset (regular MJ use prGrior to age 16; n = 24) and late onset (age 16 or later; n = 26), and 34 healthy control participants (HCs) reported smoking a minimum of 2500 times in their lives, used MJ at least five out of the last seven days, tested positive for urinary cannabinoids, and met DSM-IV criteria for MJ abuse or dependence — abstinent 12 hours
impulsivity may reflect a stable trait which precedes substance use in MJ smokers and which may predict increased MJ use. In addition, recent work has demonstrated not only that MJ smokers reported higher levels of impulsivity than non-MJ smoking individuals, but that within early onset MJ smokers (regular use prior to age 16), higher levels of impulsivity were correlated with lower levels of white matter organization and coherence (Gruber et al., 2013). This suggests that MJ is related to both impulsivity and, particularly in early onset MJ smokers, alterations of white matter, critical for efficient communication between brain regions
Participants were excluded if they met criteria for any Diagnostic and Statistical Manual of Mental Disorders (DSM-IV-TR) Axis I pathology (with the exception of MJ abuse/dependence in the smoking group)
Across the study groups, subjects were well-matched for age and IQ (123.15 in HCs, 117.56 in MJ)
No significant differences between HCs and MJ smoking groups for days of alcohol used (5.19 days of alcohol use in HC, 7.25 in MJ)
MJ per week in early onset/late onset: 24.51 smokes, 14.57 grams over 9.5 years, compared to 13.53 smokes, 5.64 grams, over 6.69 years
scores on the BIS-11 indicated significant differences between HCs and MJ smokers on all subscales (Attention, Motor, and Non-Planning) as well as Total BIS-11 scores (Table 1), demonstrating higher levels of impulsivity in MJ smokers
MJ smokers performed significantly more poorly relative to HCs during the Interference condition of the task, despite achieving similar percent accuracy during the Color Naming and Word Reading subtests
while HCs demonstrated robust posterior activation in the right cingulate cortex, MJ smokers exhibited more anterior and diffuse activation throughout the cingulate, including bilateral cingulate regions. Interestingly, results also indicated that differences between HCs and MJ smokers were primarily attributable to the early onset MJ smokers who exhibited activation in the left anterior cingulate adjacent to the genu of the corpus callosum. Late onset smokers, however, demonstrated an activation pattern more similar to the HC group, with posterior right-hemisphere activation localized to the right cingulate cortex
increased MJ use appears to represent a trait characteristic specific to the early onset MJ smokers
no between-group task differences were detected between the late MJ onset and HCs
“late onset” = 17.58 years
Sales-Carbonell C, Rueda-Orozco P, Soria-Gomez E, Buzsaki G, Marsicano G, Robbe D. 2013. “Striatal GABAergic and cortical glutamatergic neurons mediate contrasting effects of cannabinoids on cortical network synchrony” PNAS 110(2): 719-724
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3545808/ in dropbox
dual pro- and anti oscillatory functions of CB1R expressed in, respectively, subcortical and cortical regions
Samaha A-N, Seeman P, Stewart J, Rajabi H, Kapur S. 2007. “‘Breakthrough’ dopamine supersensitivity during ongoing antipsychotic treatment leads to treatment failure over time” Neuroscience 27(11): 2979-2986
http://www.jneurosci.org/content/27/11/2979.full.pdf
antipsychotic failure was accompanied by opposing changes across the synapse: tolerance to the ability of haloperidol to increase basal dopamine and dopamine turnover on one side, and 20 – 40% increases in D2 receptor number and 100 – 160% increases in the proportion of D2 receptors in the high-affinity state for dopamine (D2 High) on the other
Schizophrenic patients remain on antipsychotic (i.e., antidopaminergic) treatment for years, yet remarkably little is known about what happens to dopamine (DA) function during ongoing treatment. What is known, however, is that in both humans and laboratory animals withdrawal from chronic antipsychotic treatment reveals a state of dopaminergic supersensitivity that is characterized by increased vulnerability to psychosis and to the psychomotor activating effects of dopamine agonists, respectively
dopamine supersensitivity is not just evident on withdrawal, but develops early during antipsychotic exposure and significantly undermines the efficacy of ongoing treatment
ven when medication is guaranteed by depot injection, the average relapse rate at 1–2 years is still 18 –55% (Carpenter et al., 1999; Schooler, 2003; De Graeve et al., 2005)
Sánchez-Blázquez P, Rodríguez-Muñoz M, Garzón J.2014. “The cannabinoid receptor 1 associates with NMDA receptors to produce glutamatergic hypofunction: implications in psychosis and schizophrenia” Front Pharmacol 4: 169
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3877778/
While cannabis use in the general population does not affect the incidence of schizophrenia it does reduce the age of onset of psychotic illness among vulnerable individuals, in whom the endogenous cannabinoid system would otherwise promote the onset of this condition at a slower rate
targeted deletion of the CNR2 gene causes schizophrenia-related behaviors in mice (Ortega-Alvaro et al., 2011). This genetic finding is also supported by the results of pharmacologic experiments that were conducted in a mouse model of schizophrenia; the CB2 antagonist AM630 enhanced MK801- and methamphetamine-induced disturbances of the prepulse inhibition (PPI; Ishiguro et al., 2010) The authors concluded that reductions in CB2 function alone do not cause schizophrenia, but may be harmful for schizophrenia-susceptible individuals. Similarly, CB2 agonists reduced stereotypic behaviors that are related to schizophrenia-associated hyperdopaminergic functioning (Onaivi et al., 2006). These results are in agreement with the observation that CB2 agonists provide neuroprotection against 6-hydroxydopamine toxicity in vivo and in vitro (Garcia-Arencibia et al., 2007)
endocannabinoid system is probably the endogenous regulator that most effectively restrains NMDAR activity reducing calcium influx and the subsequent release of calcium and zinc from endogenous stores (Sensi and Jeng, 2004; Zhuang et al., 2005; Hampson et al., 2011; Sánchez-Blázquez et al., 2013b)
neuroprotective antioxidant effects of cannabinoids are also dependent upon the inhibition of Ca2+influx through voltage-gated Ca2+ channels (Mackie and Hille, 1992) and the suppression of cyclic adenosine monophosphate (cAMP)-mediated protein kinase A (PKA) signaling (Kim et al., 2006)
an endocannabinoid system that is permissive of the glutamate activity could eventually cause excitotoxic stress-induced neural dysfunctions. Excessive cannabinoid-mediated restriction of NMDAR function may precipitate glutamate hypofunction such as that observed in schizophrenia
cannabinoids that regulate NMDAR function promote the internalization/recycling of the cannabinoid receptor to reduce the stability of the glutamate receptor through the co-internalization of NR1 subunits (Garzón et al., 2009; Sánchez-Blázquez et al., 2013b). This mechanism disassembles the NMDAR and increases the presence of NR1 subunits in the cytosol, subsequently increasing the presence of NR2 subunits as well (Fan et al., 2010; Sánchez-Blázquez et al., 2013b)
As the agonist-internalized CB1s return to the cell surface within a few minutes (Hsieh et al., 1999), the presence of exogenous cannabinoids in the receptor environment produces rapid and repeated recycling of CB1s, which may cause instability and the disruption of many NMDAR
it remains unclear as to whether a safe switch disconnects NMDAR function from the negative influence that the disproportionate activation of the CB1 produces in such circumstances.
agonists that promote strong CB1s internalization, such as WIN55,212-2 and ACEA (Garzón et al., 2009), provide more effective protection against neural injury than anandamide or methanandamide which barely internalize this receptor (Kim et al., 2006; Kreutz et al., 2007)
a poor or excessive CB1-mediated effect on NMDAR activation may cause a series of neural dysfunctions in the long term
alterations in the NR1 subunits, CB1 and/or the HINT1 protein on serine/threonine kinases implicated in CB1 recruitment to NR1 subunits may cause the vulnerability required for exogenous cannabinoids to precipitate psychosis
an anomalous HINT1 protein or alterations of its endogenous regulators could contribute to cannabinoid-induced NMDAR hypofunction. The diminished expression of HINT1 mRNA found in post-mortem brains of schizophrenia patients may reflect a compensatory mechanism leading to reduce the expression of such anomalous HINT1 proteins. Moreover, data from association and expression studies suggest that variants of HINT1 may be associated with schizophrenia and that these associations may be sex-specific (Chen et al., 2008)
dysregulation of the molecular mechanisms responsible for CB1-mediated control of NMDAR activity may constitute risk factors that could precipitate symptoms of schizophrenia in early cannabis abusers that may otherwise be induced by the endogenous system, albeit at a later time
Sánchez-Blázquez P, Rodríguez-Muñoz M, Vicente-Sánchez A, Garzón J. 2013. “Cannabinoid receptors couple to NMDA receptors to reduce the production of NO and the mobilization of zinc induced by glutamate” Antioxid Redux Signal 19(15): 1766-1782
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3837442/?report=classic
NMDAR antagonism reduced the analgesia promoted by the CNR1 agonist
Cannabinoids require the HINT1 protein to counteract the toxic effects of NMDAR-mediated NO production and zinc release
Sansone R, Sansone L. 2011. “Gender Patterns in Borderline Personality Disorder” Innov Clin Neurosci 8(5): 16-20
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3115767/
Santello M, Bezzi P, Volterra A. 2011. “TNFα controls glutamatergic gliotransmission in the hippocampal dentate gyrus” Neuron 69(5): 988-1001
http://www.sciencedirect.com/science/article/pii/S0896627311001000
Glutamatergic gliotransmission provides a stimulatory input to excitatory synapses in the hippocampal dentate gyrus. Here, we show that tumor necrosis factor-alpha (TNFα) critically controls this process. With constitutive TNFα present, activation of astrocyte P2Y1 receptors induces localized [Ca2+]i elevations followed by glutamate release and presynaptic NMDA receptor-dependent synaptic potentiation. In preparations lacking TNFα, astrocytes respond with identical [Ca2+]i elevations but fail to induce neuromodulation. We find that TNFα specifically controls the glutamate release step of gliotransmission. In cultured astrocytes lacking TNFα glutamate exocytosis is dramatically slowed down due to altered vesicle docking. Addition of low picomolar TNFα promptly reconstitutes both normal exocytosis in culture and gliotransmission in situ. Alternatively, gliotransmission can be re-established without adding TNFα, by limiting glutamate uptake, which compensates slower release. These findings demonstrate that gliotransmission and its synaptic effects are controlled not only by astrocyte [Ca2+]i elevations but also by permissive/homeostatic factors like TNFα
The cytokine, released from astrocytes, was reported to strengthen excitatory synaptic transmission by promoting surface insertion of AMPA receptor (AMPAR) subunits (Bains and Oliet, 2007, Beattie et al., 2002 and Stellwagen et al., 2005)
Physiological processes like sleep have been proposed to be regulated by local variations in TNFα levels in the brain related to the sleep-wake cycle, and sleep deregulation can be induced by injections of the cytokine (Imeri and Opp, 2009 and Krueger, 2008). Therefore, an intriguing hypothesis is that the TNFα control of gliotransmission is involved in sleep homeostasis together with other glial pathways already identified (Fellin et al., 2009 and Halassa et al., 2009). Moreover, the levels of TNFα are subject to dramatic changes in pathological conditions when microglia releases large amounts of the cytokine. We have already shown in a cell culture model that in such a situation, TNFα strongly amplifies glutamate release from astrocytes (Bezzi et al., 2001). We can then hypothesize that a pathology-induced switch in the TNFα levels may have an important impact on the astrocytic input to synapses, notably in the presynaptic regulation of neuronal activity in the PP-GC hippocampal synaptic circuit. This may perturb the normal control by this circuit on critical processes such as memory formation and physiological limbic system excitability
Sapolsky R, Uno H, Rebert C, Finch C. 1990. “Hippocampal damage associated with prolonged glucocorticoid exposure in primates” Journal of Neuroscience 10(9): 2897-2902
http://www.jneurosci.org/content/10/9/2897.long in dropbox
Sar V, Akyuz G, Kugu N, Ozturk E, Ertem-Vehid H. 2006. “Axis I dissociative disorder comorbidity in borderline personality disorder and reports of childhood trauma” J Clin Psychiatry 67(10): 1583-90
http://www.ncbi.nlm.nih.gov/pubmed/17107251/
1301 college students were screened for borderline personality disorder .. 80 students with a diagnosis of borderline personality disorder and 111 nonborderline students evaluated for Dissociative Disorders by an interviewer blind to the diagnosis and scores obtained during the first phase
The prevalence of borderline personality disorder was 8.5%. A significant majority (72.5%; 58/80) of the borderline personality disorder group had a dissociative disorder, whereas this rate was only 18.0% (20/111) for the comparison group (p < .001). Childhood emotional and sexual abuse, physical neglect, and total childhood trauma scores had significant effect for borderline personality disorder (p < .001, p = .038, p = .044, and p = .003, respectively), whereas emotional neglect and diminished minimization of childhood trauma had significant effect for dissociative disorder (p = .020 and p = .007, respectively)
Sar V. 2011. “Developmental trauma, complex PTSD, and the current proposal of DSM-5” Eur J Psychotraumatol 2: 10.3402/ejpt.v2i0.5622
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3402152/
While patients with Complex PTSD share early life stress in the form of developmental traumata as a common etiological feature, nevertheless, they can also be diagnosed as having somatoform, dissociative, mood, eating, substance use, and/or borderline personality disorders depending on features of the index episode or even concurrently
nosological fragmentation leads to serious treatment impasses due to the unitary psychopathological process of individuals affected by chronic traumatization in early years of life
childhood cumulative trauma but not adult trauma predicts increasing symptom complexity in adults and cumulative trauma predicts increasing symptom complexity among children
Early research on the subject revealed that there were seven problem areas associated with early interpersonal trauma (Herman, 1992): alterations in the regulation of affective impulses, attention and consciousness, self-perception, perception of the perpetrator, systems of meaning, and somatization and/or medical problems. Accordingly, the DSM-IV described symptoms of DESNOS (Complex PTSD) as follows:
[I]mpaired affect modulation, self-destructive and impulsive behavior, dissociative symptoms, somatic complaints, feelings of ineffectiveness, shame, despair, or hopelessness, feeling permanently damaged, a loss of previously sustained beliefs, hostility, social withdrawal, feeling constantly threatened, impaired relationships with others, or a change from the individual’s previous personality characteristics
Dissociation is characterized by a disruption of usually integrated functions of memory, consciousness, identity, or perception of the environment… may also affect somatosensory functions… one of the consequences of developmental trauma
Besides constituting a disorder on its own, dissociation may accompany several psychiatric disorders including PTSD. When it does, concurrent dissociation is usually linked to a history of chronic developmental trauma in any psychiatric disorder (Sar & Ross, 2006).
Not only scientific progress, but individual patients may suffer in the medical system due to fragmented perception of consequences of human stress and would benefit from such a cohesive approach
In clinical practice it is well known that the clinical consequences of developmental trauma show themselves as the most difficult and resistant problems patients present with to mental health delivery systems. Rather than being a clue of true treatment-resistance, this common and universal phenomenon may be caused by inability of mental health delivery systems to accurately diagnose and address the needs of these patients.
Savitz J, van der Merwe L, Stein D, Solms M, Ramear R. 2007. “Genotype and childhood sexual trauma moderate neurocognitive performance: a possible role for brain-derived neurotrophic factor and apolipoprotein E variants” Biol Psychiatry 62(5): 391-9
http://www.ncbi.nlm.nih.gov/pubmed/17210134
sexual abuse produces adverse effect on memory performance; gene-environment interaction
Schacht J, Hutchison K, Filbey F. 2012. “Associations between Cannabinoid Receptor-I (CNRI) Variation and Hippocampus and Amygdala Volumes in Heavy Cannabis Users” Neuropsychopharmacology 2012: 37; 2368-2376
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3442352/ in dropbox
all CU reported significantly more alcohol use (drinks/drinking day and drinking days/month) than HC, more likely to be cigarette smokers
Controlling for total intracranial volume and other confounding variables, matched cannabis users had smaller bilateral hippocampi and left amygdalae than controls
On average, left and right hippocampi were 6.9 and 7.1% smaller, respectively, and left amygdala was 5.8% smaller among CU than among HC. difference in right amygdalar volume between groups (5.6% smaller among CU) not significant
a common genetic variant may moderate volume differences in medial temporal structures related to heavy cannabis use (difference in G allele CU/HC, but not in A allele)
interaction between rs202329 genotype and group on hippocampal volume indicates G allele engendered smaller hippocampal volume only among CU; among HC, G-allele carriers had larger bilateral hippocampi than A-allele carriers
it is also possible that smaller hippocampal volume and rs2023239 genotype independently preceded heavy cannabis use in the current study
associations between rs202329 variance and cannabis withdrawal, craving, and cue-elicited brain activation (Filbey et al, 2010; Haughey et al, 2008), alcohol cue-elicited brain activation and greater subjective reward after consuming alcohol (Hutchison et al, 2008), greater alcohol craving at higher levels of alcohol consumption (van den Wildenberg et al, 2007), greater trait impulsivity (Ehlers et al, 2007), greater severity of nicotine dependence (Chen et al, 2008).
The HC sample, and particularly the subset of HC with an rs2023239 G allele (N=12), was relatively small, and outliers might have contributed to the magnitude of the genotype by group interactions.
Schaeffer J, Andrysiak T, Ungerleider J. 1981. “Cognition and long-term use of ganja (Cannabis)” Science 213(4506): 465-6
http://www.ncbi.nlm.nih.gov/pubmed/6972600/
No impairment of cognitive function was found
Schafe G, Doyere V, LeDoux J. 2005. “Tracking the fear engram: the lateral amygdala is an essential locus of fear memory storage” Journal of Neuroscience 25(43): 10010-10014
http://www.jneurosci.org/content/25/43/10010.full
auditory fear conditioning induces alterations in the activity of neurons not only in the lateral amygdala but also in the auditory cortex (Bakin and Weinberger, 1990; Edeline and Weinberger 1993) and the auditory thalamus (Gabriel et al., 1975;Lennartz and Weinberger, 1992)
Collectively, our findings show that long-term synaptic plasticity in the LA is required for both memory consolidation of fear conditioning and the consolidation of training-induced plasticity in the LA
Scheller-Gilkey G, Moynes K, Cooper I, Kant C, Miller A. 2004. “Early life stress and PTSD symptoms in patients with comorbid schizophrenia and substance abuse” Schizophr Res 69(2-3): 167-74
http://www.ncbi.nlm.nih.gov/pubmed/15469190
Patients with schizophrenia and a history of substance abuse had significantly higher frequency and severity of childhood traumatic events, greater PTSD symptomatology, significantly higher depression scores, and higher scores on the general subscale of the PANSS.
Schmid-Schönbein G. 2009. “Inflammation and the autodigestion hypothesis” Microcirculation 16(4): 289-306
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2677689/
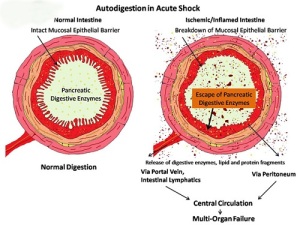
Systemic inflammatory cascade can damage many cell types, including the protective layers responsible for containing pancreatic digestion enzymes; this cascade can therefore allow these enzymes to enter circulation, leading to autodigestion (image from Schmid-Schönbein, 2009)
Schmid-Schönbein G, Chang M. 2014. “The autodigestion hypothesis for shock and multi-organ failure” Ann Biomed Eng 42(2): 405-414
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3943906/
Shock is accompanied by strong markers for inflammation and involves a cascade of events that leads to failure in organs even if they are not involved in the initial insult… pancreatic digestive enzymes carried in the small intestine after mixing with ingested food are a major cause for multi-organ failure
Schreurs B, Burhans L. 2015. “Eyeblink classical conditioning and post-traumatic stress disorder — a model systems approach” Front Psychiatry 6: 50
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4389289/
People exposed to trauma who suffer flashbacks, bad dreams, numbing, fear, anxiety, sleeplessness, hyper-vigilance, hyperarousal, or an inability to cope comprise the 15–25% who suffer from post-traumatic stress disorder (PTSD) (1–3)
the majority of animal models rely on group data, and it is clear that not everyone exposed to trauma develops PTSD (2, 13, 30, 34, 35). In fact, depending on the population and on the type of trauma, only 5–25% of exposed people develop PTSD (1–3)
The consensus of these studies is that there may be changes in EBCC as a result of PTSD but the effects are quite variable and may involve personality traits (57)
clinical data in which exacerbation or reactivation of prior symptoms accounts for 38.3% of military cases of PTSD and 15.3% of civilian cases (120, 125)
Schuff N, Zhang Y, Zhan W, Lenoci M, Ching C, Boreta L, Mueller S, Wang Z, Marmar C, Weiner M, Neylan T. 2011. “Patterns of altered cortical perfusion and diminished subcortical integrity in posttraumatic stress disorder: an MRI study” Neuroimage 54 suppl 1: S62-8
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2945438/
Seventeen male veterans with PTSD (45 ± 14 years old) and 15 age-matched male veterans without PTSD
PTSD was associated with a smaller dentate gyrus and a smaller CA3 region, which is a major target for glucocorticoids
DTI studies suggested that PTSD is associated with microstructural degradation of the cingulum bundle
right parietal lobule showed systematically elevated rCBF values in PTSD right parietal cortex (Brodmann Area), right superior temporal cortex, right inferior frontal gyrus and right superior frontal gyrus
PTSD had reduced FA in white matter regions of the prefrontal lobe, incl areas near the anterior cingulate cortex (ACC), prefrontal cortex (PFC) and precentral gyrus (PCG), as well as in the posterior internal capsule (p.IC) and the posterior angular gyrus (AG)
studies using electroencephalography also found a unique pattern of tonically high right parietal and low left frontal activation in PTSD, (Metzger et al., 2004) more right than left activation during anxious arousal (Nitschke et al., 1999)
while we found increased rCBF in the inferior parietal lobule in PTSD, several functional studies reported decreased rCBF in this region when PTSD patients were confronted with provocative trauma stimuli
compromised function of these regions in PTSD (for a review see (Lanius et al., 2006)) as well as with MRI findings of reduced size (Woodward et al., 2006; Yamasue et al., 2003) and metabolism (Schuff et al., 2008) of the anterior cingulate cortex in PTSD
Studies on rats showed that stress can lead to a decrease in the number of dendritic length and branch points in the anterior cingulate (Radley et al., 2006)
repeated stress activates the glutamate neurotransmission in the anterior cingulate that leads in turn to persistent activation of extracellular-regulated kinase, causing neuronal damage by destabilizing the cytoskeletal (for a review see (Moghaddam, 2002))
disruption of anterior cingulate and prefrontal cortex function is integral to many symptoms of PTSD due to impaired extinction of fear conditioning
functional neuroanatomic model of PTSD which posits hyperactivation of the amygdala and insula in response to external stressors together with hypoactivation of anterior structures particularly the anterior cingulate cortex and medial prefrontal cortex (Etkin and Wager, 2007)
inferior temporal lobe is difficult to cover with ASL. It is therefore possible that we missed important physiological alterations in this region
Schuitevoerder S, Rosen J, Twamley E, Ayers C, Sones H, Lohr J, Goetter E, Fonzo G, Holloway K, Thorp S. 2013. “A meta-analysis of cognitive functioning in older adults with PTSD” Journal of Anxiety Disorders 2013: 27; 550-558
http://www.ncbi.nlm.nih.gov/pubmed/23422492 in dropbox
– sample is almost entirely holocaust survivors; remainder combat veterans
worse performance across cognitive measures in older adult samples with PTSD relative to older samples without PTSD. strongest effect in memory,
The effect of learning was about half the magnitude for comparison subjects of PTSD+ vs. PTSD− as the PTSD+ vs. HC, trauma exposure may uniquely contribute to worse performance in learning
Schultz C, Nenadic I, Koch K, Wagner G, Roebel M, Schachtzabel C, Muhleisen W, Nothen M, Cichon S, Deufel T, Kiehntopf M, Rietschel M, Reichenbach J, Sauer H, Schlosser R. 2011. “Reduced cortical thickness is associated with the glutamatergic regulatory gene risk variant DAOA Arg30Lys in schizophrenia” Neuropsychopharmacology 36(8): 1747-1753
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3138664/
cortical thinning in theory of mind network and visual processing linked with glutamatergic function
Schweinsburg A, Brown S, Tapert S. 2008. “The influence of marijuana use on neurocognitive functioing in adolescents” Curr Drug Abuse Rev 1(1): 99-111
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2825218/
Marijuana use is common in adolescence, yet neural consequences have not been well delineated
the impact of preexisting risk factors is unknown
Adolescents demonstrate persisting deficits related to heavy marijuana use for at least six weeks following discontinuation, particularly in the domains of learning, memory, and working memory. Further, adolescents appear more adversely affected by heavy use than adults. Longitudinal studies will help ascertain whether preexisting differences contribute to these abnormalities
among adults with a lifetime marijuana use disorder (DSM-IV criteria for abuse or dependence), 82% also met criteria for an alcohol use disorder, and 48% met criteria for nicotine dependence [3]
individuals in treatment manifest more severe substance use disorders, and poorer social, cognitive, and behavioral functioning [4] (than users not seeking treatment)
Adults who use marijuana chronically have demonstrated poorer performance on tests of learning and memory, attention, visuospatial skills, processing speed, and executive functions [5–11]. However, some investigations have found no performance decrements among heavy cannabis users [12–15]. A meta-analysis examined 11 studies that met strict inclusion criteria, and ascribed impaired learning and memory to chronic marijuana consumption, but determined that other cognitive domains remain unaffected [16]
Within 6 – 36 hours following use, marijuana users showed increased and widespread spatial working memory activation, both in anterior cingulate and prefrontal regions normally associated with spatial working memory, as well as in additionally recruited brain areas that were not activated among controls [27]
During inhibitory processing, heavy marijuana users evidenced reduced anterior cingulate and greater mid-cingulate response, as well as more widespread bilateral dorsolateral prefrontal activation [29]
Marijuana users demonstrated poorer verbal recall after a 25 minute delay than comparison youths during nicotine withdrawal, but not during the smoking condition. On the verbal working memory task, marijuana users showed a greater reduction in accuracy with increasing working memory load relative to comparison teens – Jacobsen
Neurocognitive abnormalities among marijuana users were observed during nicotine withdrawal but not during ad libitum smoking, indicating that nicotine use may mask the deficits associated with marijuana use
Despite similar task performance, marijuana users exhibited increased fMRI response during both inhibitory and non-inhibitory trials of the go/no-go task, particularly in dorsolateral prefrontal and parietal regions. Those with a later age of onset and briefer duration of regular use showed the greatest increase in brain response
Recent users showed more fMRI response compared to abstinent users in medial and left superior prefrontal cortices, bilateral insula, and right superior parietal cortex, suggesting increased reliance on working memory updating, spatial rehearsal strategies, and inhibitory control
continuous measures of abstinence duration were not significantly associated with levels of neurocognitive performance
After reviewing the literature, there is preliminary evidence of persisting neurocognitive abnormalities among adolescent marijuana users. Subtle deficits in learning and memory, working memory, and attention
it is still unknown whether marijuana use caused or contributed to these effects. In most cases, decrements were observed among those with relatively heavy, regular marijuana use. Lighter use, even if chronic, was often not associated with neurocognitive dysfunction. Thus, it is unclear whether only very heavy use is detrimental to brain functioning, or whether very heavy users differ from lighter users on other factors that account for abnormalities
preexisting factors that weren’t measured may have contributed to cognitive differences between groups. For instance, the types of youths who initiate heavy marijuana use at a young age may perform more poorly in school because of social, cognitive, and motivational factors that continue to affect social and occupational attainment into adulthood, which in turn may contribute to neurocognitive differences. Moreover, both animal and human studies have indicated an association between heavy marijuana use and behavioral features such as increased anxiety, affective, and psychotic symptoms [81–85], particularly among those who may have pre-existing vulnerabilities [84]. Interactions between genes, the environment, and such behavioral factors may influence marijuana use and neurocognition. Although beyond the scope of this review, future studies should attempt to identify the possible premorbid features that contribute to neurocognitive abnormalities among adolescent marijuana users
Schweinsburg A, Schweinsburg B, Medina K, McQueeny T, Brown S, Tapert S. 2010. “The influence of recency of use on fMRI response during spatial working memory in adolescent marijuana users” J Psychoactive Drugs 42(3): 401-412
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3016644/
Participants were 13 recent users (2 – 7 days abstinent), 13 abstinent users (27 – 60 days abstinent), and 18 non-using controls, all ages 15 – 18. Groups were similar on demographics, had no psychiatric or medical disorders, and user groups were similar on substance histories
Groups performed similarly on the task, but recent users showed greater fMRI response in medial and left superior prefrontal cortices, as well as bilateral insula. Abstinent users had increased response in the right precentral gyrus (clusters ≥1328 μl, p<.05).
Most of the marijuana users in this study were moderate to heavy drinkers. Our previous research characterized neural response differences between adolescent alcohol and marijuana users compared to users of alcohol alone
Anxiety levels were correlated with brain response in the current study
Scott J, Chant D, Andrews G, Martin G, McGrath J. 2007. “Association between trauma exposure and delusional experiences in a large community-based sample” Br J Psychiatry 190: 339-43
http://www.ncbi.nlm.nih.gov/pubmed/17401041
(n=10 641) Exposure to any traumatic event but without the development of PTSD was associated with increased endorsement of delusional experiences (relative risk 2.68, 95% CI 2.18-3.30) and there was a significant dose-response relationship between the number of types of traumatic events and endorsement of such experiences (chi(2) =26.74, d.f.=2, P<0.001). A diagnosis of PTSD further increased endorsement of delusional experiences (RR=9.24, 95% CI 6.95-12.27). The association between PTSD and delusional experiences remained significant after adjusting for factors associated with psychotic symptoms
Sexton C, Mackay C, Ebmeier K. 2009. “A systematic review of diffusion tensor imaging studies in affective disorders” Biol Psychiatry 66: 814-823
http://www.ncbi.nlm.nih.gov/pubmed/19615671 in dropbox
Shanks N, Windle R, Perks P, Harbuz M, Jessop D, Ingram C, Lightman S. 2000. “Early-life exposure to endotoxin alters hypothalamic-pituitary-adrenal function and predisposition to inflammation” Proc Natl Acad Sci USA 97(10): 5645-5650
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC25882/
Exposure of neonatal rats to a low dose of endotoxin resulted in long-term changes in hypothalamic–pituitary–adrenal (HPA) axis activity, with elevated mean plasma corticosterone concentrations that resulted from increased corticosterone pulse frequency and pulse amplitude
neonatal endotoxin exposure had long-lasting effects on immune regulation, including increased sensitivity of lymphocytes to stress-induced suppression of proliferation and a remarkable protection from adjuvant-induced arthritis
a potent and long-term effect of neonatal exposure to inflammatory stimuli that can program major changes in the development of both neuroendocrine and immunological regulatory mechanisms
activation of endocrine and immune systems during neonatal development can program or “reset” functional development of both the endocrine and immune systems
Sheline Y, Gado M, Price J. 1998. “Amygdala core nuclei volumes are decreased in recurrent major depression.” Neuroreport 9(9):2023-8
http://www.ncbi.nlm.nih.gov/pubmed/9674587
20 patients with a history of depression and 20 pair-wise matched controls.
Depressed subjects had bilaterally reduced amygdala core nuclei volumes and no significant differences in total amygdala volumes or in whole brain volumes. Since patients with a depression history have bilateral hippocampal volume reduction the volume loss in this closely related structure suggests a shared effect on both structures, potentially glucocorticoid-induced neurotoxicity mediated by the extensive reciprocal glutamatergic connections
Shevlin M, Dorahy M, Adamson G. 2007. “Childhood traumas and hallucinations: an analysis of the National Comorbidity Survey” J Psychiatr Res 41(3-4):222-8
http://www.ncbi.nlm.nih.gov/pubmed/16643948
a history of childhood rape and molestation were significantly associated with visual, auditory and tactile hallucinations. Additionally, neglect was associated with visual hallucinations and physical abuse with tactile hallucinations. Experiencing multiple types of trauma was associated with increases in the likelihood of reporting each of the three types of hallucinations. Hallucinatory experiences are possible indicators of a traumatic childhood history
Shiflett M, Rankin A, Tomaszycki M, DeVoogd T. 2004. “Cannabinoid inhibition improves memory in food-storing birds, but with a cost” Proc Biol Sci 271(1552):2043-8
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1691822/
Intra-hippocampal infusions of the CB1-R antagonist SR141716A enhanced long-term memory for the location of a hidden food reward, measured 72 h after encoding. However, when the reward location changed during the retention interval, birds that had received SR141716A during initial learning showed impairments in recalling the most recent reward location. Thus, blocking CB1-R activity may lead to more robust, long-lasting memories, but these memories may be a source of proactive interference
Shin L, Orr S, Carson M, Rauch S, Macklin M, Lasko N, Peters P, Metzger L, Dougherty D, Cannistraro P, Alpert N, Fischman A, Pitman R. 2004. “Regional cerebral blood flow in the amygdala and medial prefrontal cortex during traumatic imagery in male and female Vietnam veterans with PTSD” Arch Gen Psychiatry 61(2): 168-76
http://www.ncbi.nlm.nih.gov/pubmed/14757593/
symptom severity was positively related to rCBF in the right amygdala and negatively related to rCBF in medial frontal gyrus.
suggest a reciprocal relationship between medial prefrontal cortex and amygdala function in PTSD and opposing associations between activity in these regions and symptom severity
Shoemaker J, Ruckle M, Mayeux P, Prather P. 2005. “Agonist-directed trafficking of response by endocannabinoids acting at CB2 receptors” JPET 315(2): 828-838
http://jpet.aspetjournals.org/content/315/2/828.long
three intracellular effectors via CB2 receptors in transfected Chinese hamster ovary cells. Although the three agonists regulate all effectors with equivalent efficacy, the rank order of potencies differs depending on which effector is evaluated
endocannabinoids also seem to be more “efficient” agonists at CB2 receptors relative to synthetic agonists. 2-AG and noladin ether require occupancy of less than one-half the number of receptors to produce comparable regulation of adenylyl cyclase and ERK-MAPK, relative to the synthetic cannabinoid CP-55,940
Numerous studies provide support that individual agonists acting at several different classes of GPCRs (Figini et al., 1997; Berg et al., 1998; Wiens et al., 1998), including CB1 receptors (Bonhaus et al., 1998), are able to traffic intracellular responses in a ligand-dependent manner
Shonkoff J, Boyce W, McEwen B. 2009. “Neuroscience, Molecular Biology, and the Childhood Roots of Health Disparities Building a New Framework for Health Promotion and Disease Prevention.” JAMA 301(21): 2252-2259
http://jama.jamanetwork.com/article.aspx?doi=10.1001/jama.2009.754
there can be a lag of many years, even decades, before early adverse experiences are expressed in the form of disease
confronting origins of disparities may produce greater effects than attempting to modify health-related behaviours in adulthood
Sidney S, Quesenberry C Jr, Friedman G, Tekawa I. 1997. “Marijuana use and cancer incidence (California, United States)” Cancer Causes Control 8(5): 722-8
http://www.ncbi.nlm.nih.gov/pubmed/9328194/
The study population consisted of 64,855 examinees in the Kaiser Permanente multiphasic health checkup in San Francisco and Oakland (California, United States), between 1979-85, aged 15 to 49 years
Compared with nonusers/experimenters (lifetime use of less than seven times), ever- and current use of marijuana were not associated with increased risk of cancer of all sites (relative risk [RR] = 0.9, 95 percent confidence interval [CI] = 0.7-12 for ever-use in men; RR = 1.0, CI = 0.8-1.1 in women) in analyses adjusted for sociodemographic factors, cigarette smoking, and alcohol use. Marijuana use also was not associated with tobacco-related cancers or with cancer of the following sites: colorectal, lung, melanoma, prostate, breast, cervix. Among nonsmokers of tobacco cigarettes, ever having used marijuana was associated with increased risk of prostate cancer (RR = 3.1, CI = 1.0-9.5) and nearly significantly increased risk of cervical cancer (RR = 1.4, CI = 1.0-2.1)
Silins E, Horwood L, Patton G, Fergusson D, Olsson C, Hutchinson D, Spry E, Toumbourou J, Degenhardt L, Swift W, Coffey C, Tait R, Letcher P, Copeland J, Mattick R. 2014. “Young adult sequelae of adolescent cannabis use: an integrative analysis” Lancet Psychiatry 2014 1: 286-93
http://www.thelancet.com/journals/lanpsy/article/PIIS2215-0366(14)70307-4/abstract in dropbox
association with depression disappeared after adjusting for confounds. Low educational attainment, high drug use and suicide risk remained significant.
Simon A, Gorman J. 2006. “Advances in the treatment of anxiety: targeting glutamate” NeuroRx 3(1): 57-68
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3593356/
Our current psychopharmacological treatments for anxiety disorders evince a number of shortcomings, including troublesome side effects and lack of primary effects
In normal human subjects, fear and anxiety are produced in response to a variety of exteroceptive, interoceptive, and cognitive inputs and allow the individual to assess his or her environment and make appropriate behavioral adjustments. In life-threatening situations, the brain’s anxiety network allows for this activation below the level of conscious awareness such that responses can occur as quickly as possible
post-traumatic stress disorder (traumatic cues), panic disorder (bodily sensations), social phobia (interpersonal ridicule/embarrassment), and specific phobias (specific items)
we cannot assert that any of our current treatments actually target a known pathophysiology related to anxiety, frustrating our ability to make progress in developing more effective medications
the field of developing pharmacological treatments for mood and anxiety disorders is currently mired in a reiterative process that persists in coming up with variations on the monoamine reuptake inhibitor strategy—a gross intervention discovered by chance
one unifying pathophysiological feature of several anxiety conditions is excessive excitatory amino acid neurotransmission in response to stress
1) exaggerated response to stress due to constitutive and genetic factors27,42,43 and/or 2) exposure to chronic levels of stress, are key components in the vulnerability to develop mood and anxiety disorders
Glutamate receptors are split into several types, most broadly demarcated as ionotropic and metabotropic.52 The former include the NMDA, AMPA, and kainate receptors. The metabotropic receptors have a modulatory role
dysregulated and excessive activation leads to excitotoxicity with increased calcium ion entry and death of neurons
A neutral stimulus leads to a certain amount of amygdalar glutamate release that binds to NMDA and AMPA/kainate receptors, but this weak activation and depolarization are not strong enough to dislodge the Mg2 that partially blocks the NMDA channel. A strong and aversive stimulus, on the other hand, produces sufficient depolarization to allow full permeability of the NMDA receptor and ion channel. When administered together, the previously neutral stimulus begins to take on the neurochemical signature of the aversive stimulus.
acute fear in normal humans is reliably accompanied by activation of the amygdala65 and that exaggerated amygdala activity occurs in patients with depression,66 panic disorder,67 social anxiety disorder,68 posttraumatic stress disorder,69 and trait worry in individuals without a psychiatric illness
chronic, but not acute, administration of citalopram induces downregulation of the NR2B subunit of the NMDA receptor
rats that underwent restraint-tail-shock stress manifested an increase in hippocampal LTD via a mechanism by which stress activates glucocorticoid receptors, inhibiting the uptake of glutamate, leading to a spillover and temporal summation of glutamate at extrasynaptic NR2B-containing NMDA receptors, inducing hippocampal LTD… stress may lead to a phosphorylation-induced inactivation of glutamate-aspartate transporters, thus allowing the extrasynaptic spillover of glutamate
NR2B is more important than NR2A during brain development, and although this reverses as the brain matures, authors make important statements that NR2B subunits still abound in the mature brain
Bruce McEwen (see reviews by McEwen47 and Sapolsky103) originally showed evidence of hippocampal dendritic remodeling in response to stress
In the case of chronic stress, high cortisol levels in conjunction with glutamate excess could contribute to this cellular injury
one mechanism for terminating excessive glutamatergic neurotransmission and the sustained activation of NMDA receptors is the internalization of these receptors.118 Interestingly, NMDA receptors are functionally downregulated at synapses as a result of fear learning.117 We hypothesize that, during acute stress, internalization of glutamatergic receptors occurs and functions as an adaptive method of reducing the excitotoxic effects of glutamate on postsynaptic neurons, and during more excessive or prolonged stress, the neuron further adaptively reduces the number of available glutamate receptors by reducing dendrite length and spine number to protect itself from glutamate-induced cell death. Although this limits glutamatergic toxic effects on the neuron and may preserve the cell body, it essentially disconnects the neuron from other neurons by reducing the number of synaptic connections, thereby preventing cell death, but possibly precipitating anxiety or depressive disorders
by reducing the cell surface expression of NMDA receptors, these drugs may thereby reduce the sensitivity of the neuron to glutamate, and this decrease in glutamatergic neurotransmission may avert dendritic remodeling under stress conditions
eight receptors are divided into three groups (group I, mGluR1/5; group II, mGluR2/3; and group III, mGluR4/ 6/7/8)… group I receptors are primarily (though not exclusively) located postsynaptically and serve an excitatory role… mGluR5 has been postulated to be required for fear memory formation and long-term potentiation in the amygdala… group II receptors exist at both pre- and postsynaptic sites and are inhibitory. Whereas it has been shown that their presynaptic activation serves to decrease glutamate release (and that of other neurotransmitters)152 via a negative feedback mechanism, it has also been argued that postsynaptic agonism limits the excitability of target neurons
changes in group I and group II metabotropic receptor expression in the prefrontal cortex of rats exposed to an impoverished rearing environment
NAA concentration is reduced in classic degenerative brain disorders like Huntington’s disease,166 Alzheimer’s disease,167 and HIV-associated dementia.168 It has also been shown to be abnormal in key brain areas in several psychiatric illnesses including schizophrenia,169 depression,170 and anxiety disorders,4,171–173 although the directionality of NAA in these diseases has been more variable than in the neurodegenerative disorders. We and others have shown, for example, decreased NAA in the superior temporal gyrus of patients with schizophrenia,174 decreased NAA in the anterior cingulate of monkeys exposed to early rearing stress,175 reduced NAA concentrations in the temporal cortex of rats reared in isolation,176 and increased NAA in the dorsolateral prefrontal cortex of patients with GAD.4 Importantly, NAA appears to be vulnerable to psychosocial stress. Additional preliminary data from the latter study also showed that cognitive behavioral therapy decreases NAA in the dorsolateral prefrontal cortex in patients with GAD
Simons J, Carey K. 2002. “Risk and vulnerability for marijuana use problems: the role of affect dysregulation” Psychol Addict Behav 16(1):72-5
http://www.ncbi.nlm.nih.gov/pubmed/11934090
Association between lability, affect dysregulation and problem use, but not use.
Siqueira L, Diab M, Bodian C, Rolnitzky L. 2001. “The relationship of stress and coping methods to adolescent marijuana use” Subst Abus 22(3): 157-166
http://www.ncbi.nlm.nih.gov/pubmed/12466675
18 adolescents aged 12-21 years. Lifetime use in this population was 59% (n = 611) with 18.4% (n = 191) reporting frequent weekly use
negative life events, greater use of the negative coping method of anger and less frequent use of the positive coping method of parental support related to use; did not report greater stress
Skosnik P, Edwards C, O’Donnell B, Steffen A, Steinmetz J, Hetrick W. 2008. “Cannabis use disrupts eyeblink conditioning: evidence for cannabinoid modulation of cerebellar-dependent learning” Neuropsychopharmacology 33(6): 1432-1440
the cerebellum contains the highest density of cannabinoid receptor (CB1) in the brain
current study examined the effect of chronic cannabis use on classical eyeblink conditioning (EBC), a cerebellar-mediated task which has been shown to be disrupted in CB1 knockout mice. Chronic cannabis users (24 hours abstinence prior to study; positive THC urine drug test) free of DSM-IV Axis I or Axis II disorders, were evaluated
Purkinje cells activated by glutamatergic granule cells normally serve to inhibit cerebellar deep nuclei (i.e. interpositus nucleus) thus modulating the timing of conditioned blink responses (Steinmetz, 2000). Chronic cannabis use, which is known to produce behavioral tolerance and may induce long-term endocannabinoid compensatory changes (e.g. CB1 downregulation; Sim et al, 1996; Romero et al, 1998), could result in increased granule cell glutamate release, and hence overactive inhibitory Purkinje inputs to the interpositus
Current cannabis users (n = 14) and healthy drug-naive controls (n = 10) were assessed… from the local university community… avg age of first cannabis 15.3, far more more alcohol and cigarettes in cannabis group, moderately lower intelligence scores across the board, 9.7 joints per week average. 38% diagnosed with cannabis abuse, 21% with dependence
Percent CRs and CR peak latency during each 10-trial block (plus extinction) were calculated for each group. For % CRs (), a repeated measures ANOVA revealed a main effect of block [F(9,198) = 8.67, p < 0.0001; partial η2 = 0.85] and a main effect of group [F(1,22) = 14.36, p < 0.001; partial η2 = 0.40]. However, there was no group X block interaction [F(9,198) = 1.89, p = 0.14; partial η2 = 0.55]. Given that the cannabis group exhibited slightly higher alcohol use rates than controls (see ), the average number of drinks per week (during the past month) was included in the repeated measures ANOVA as a covariate. The main effect of group remained significant after covarying for alcohol use [F(1,21) = 9.63, p < 0.005; partial η2 = 0.31]. Likewise, the group differences also remained significant after covarying for tobacco use per day [F(1,21) = 12.80, p < 0.002; partial η2 = 0.38]. For CR peak latency, there was also a main effect of block [F(9,198) = 5.17, p < 0.003; partial η2 = 0.77] and a main effect of group [F(1,22) = 7.23, p < 0.01; partial η2 = 0.25]. There was also a trend towards a group X block interaction [F(9,198) = 2.31, p = 0.08; partial η2 = 0.60]. The main effect of group remained significant after covarying for tobacco use [F(1,21) = 7.85, p < 0.01; partial η2 = 0.27], and fell to a trend level difference after covarying for alcohol use [F(1,21) = 2.79, p < 0.10; partial η2 = 0.12]. For UR amplitude, no group differences were observed during paired trials [F(1,22) = 0.71, p = 0.41] or in UR latency during paired trials [F(1,22) = 0.02, p = 0.9]
extinction rates for the two groups were equal. … for the ERP measure, a one-way ANOVA revealed that there were no differences in N100 amplitude between the groups [F(1,23) = 0.11, p = 0.75]
inhibitory Purkinje cells tonically suppress interpositus activity, and it is thought that for conditioning to occur, the GABAergic Purkinje cells must themselves be inhibited, which in turn disinhibits interpositus activity, thus affecting the timing and gain of CRs (Steinmetz, 2000)
since the current study did not directly administer cannabis or THC, it remains possible that some pre-existing condition contributed to both cannabis drug-seeking and altered EBC performance (e.g. premorbid alterations in CB1 or endocannabinoids). However, since all of the current cannabis subjects tested positive for THC, and had a mean of 5.6 years of cannabis use, the informed assumption of CB1 downregulation discussed above is likely more proximal and salient than subtle pre-existing individual differences in endocannabinoid activity. Nonetheless, since there was no way of directly assessing CB1 densities in the current sample, possible preexisting differences cannot be ruled out using the current methodology
Slavich G, Thornton T, Torres L, Monroe S, Gotlib I. 2009. “Targeted rejection predicts hastened onset of major depression” J Soc Clin Psychol 28(2): 223-243
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2847269/
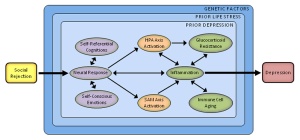
Twenty-seven adults with major depressive disorder were administered an interview-based measure of life stress. Severe life events that occurred prior to the onset of depression were subsequently coded as targeted rejection (TR) or as non-TR. Participants who experienced a pre-onset severe TR event became depressed approximately three times faster than did their non-TR counterparts
Slavich G, O’Donovan A, Epel E, Kemeny M. 2010. “Black sheep get the blues: a psychobiological model of social rejection and depression” Neurosci Biobehav Rev 35(1): 39-45
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2926175/
Major life events involving social rejection are strongly associated with onset of depression. To account for this relation, we propose a psychobiological model in which rejection-related stressors elicit a distinct and integrated set of cognitive, emotional, and biological changes that may evoke depression. In this model, social rejection events activate brain regions involved in processing negative affect and rejection-related distress (e.g., anterior insula, dorsal anterior cingulate cortex). They also elicit negative self-referential cognitions (e.g., “I’m undesirable,” “Other people don’t like me”) and related self-conscious emotions (e.g., shame, humiliation). Downstream biological consequences include upregulation of the hypothalamic-pituitary-adrenal axis, sympathetic-adrenal-medullary axis, and inflammatory response. Pro-inflammatory cytokines play an important role in this process because they induce a constellation of depressotypic behaviors called sickness behaviors. Although these changes can be short-lived, sustained inflammation may occur via glucocorticoid resistance, catecholamines, sympathetic innervation of immune organs, and immune cell aging. This response also may be moderated by several factors, including prior life stress, prior depression, and genes implicated in stress reactivity
Depressed individuals are 2.5- to 10-times more likely than nondepressed persons to have experienced a recent major life event (Kendler et al., 1995; Shrout et al., 1989), and up to 82% of depressive episodes appear to be precipitated by such stress (Mazure, 1998)
We focus on social rejection because rejection-related life events increase risk for depression to a greater degree than any other type of stress (Kendler et al., 2003)
Interpersonal loss has long been regarded as central to depression. This focus can be traced back to Freud’s (1917/1957) Mourning and Melancholia, and is represented in many contemporary psychodynamic (Blatt, 2004), cognitive (Beck, 1967), and social (Brown and Harris, 1978) accounts
individuals who break up with their partner are 10.2-times more likely to develop depression than those without such stress. However, individuals who are broken-up with are 21.6-times more likely to develop depression (Kendler et al., 2003)
Individuals who experienced a recent severe Targeted Rejection event became depressed approximately three times faster than those who experienced a severe life event that was not defined as TR
rejection events may elicit negative self-referential cognitions concerning one’s social worth and esteem, such as “I’m undesirable,” “I’m unlovable,” and “Other people don’t like me” (Monroe et al., 2007a)
shame and humiliation, which are associated with biological processes (e.g., inflammation) that support behavioral disengagement and withdrawal (Kemeny, 2009)
anterior insula and dorsal anterior cingulate cortex (dACC; Lieberman and Eisenberger, 2009). Although activation in multiple biological systems may follow, the hypothalamic-pituitary-adrenal (HPA) axis and sympathetic-adrenal-medullary (SAM) axis are particularly responsive. Degree of reactivity differs based on subtle features of the social environment, such as availability of social support (O’Donovan and Hughes, 2008). Moreover, this reactivity may be indexed by two biomarkers: the glucocorticoid cortisol, which is released by the HPA axis, and the catecholamines epinephrine and norepinephrine, which are released by the SAM axis
meta-analytic review of 208 studies revealed that rejection-related stressors are particularly activating (Dickerson and Kemeny, 2004)
cytokines are notable because they signal the central nervous system to elicit sickness behaviors. Sickness behaviors are thought to facilitate an organism’s recuperation and recovery from illness or injury, and include depressotypic symptoms such as anhedonia, fatigue, psychomotor slowing, and social-behavioral withdrawal (Dantzer et al., 2008; Yirmiya et al., 2000)
shame in everyday life, also has been associated with elevated in vitro lipopolysaccharide-stimulated levels of IL-6 (Rohleder et al., 2008)… social rejection, specifically social isolation (Cole et al., 2007) and interpersonal stress (Miller et al., 2009b), upregulate the expression of genes that promote inflammation.
laboratory stressors involving social-evaluative threat reliably elicit increases in IL-1β, IL-6, and TNF-α (Kemeny, 2009)
administration of the pro-inflammatory cytokines IL-1β and TNF-α elicits a depressotypic behavioral state in rodents (Dantzer et al., 2008). Second, administration of cytokines that promote inflammatory activity, specifically interferon-α (IFN-α) and interleukin-2 (IL-2), evokes clinically-significant levels of depression in up to 50% patients who receive them as treatments for infectious diseases or cancer (Capuron and Miller, 2004; Raison et al., 2006). Finally, vaccination and administration of endotoxin, both of which increase pro-inflammatory cytokine production, induce depressed mood in physically healthy individuals (Eisenberger et al., 2009; Glaser et al., 2003)
administration of IFN-α was found to be associated with greater dACC activation in participants completing a visuospatial attention task (Harrison et al., 2009) Also, endotoxin-induced mood deterioration has been associated with activation in the subgenual anterior cingulate cortex, a brain region implicated in mood regulation and in the etiology of depression (Capuron et al., 2005; see also O’Connor et al., 2009).
exposure to social threat promotes the production of cortisol and catecholamines. Binding of circulating cortisol and catecholamines to receptors on immune cells can modulate the release of pro-inflammatory cytokines (Padgett and Glaser, 2003). Typically, cortisol acts on the glucocorticoid receptor with anti-inflammatory effects (Sapolsky et al., 2000). Prolonged exposure to cortisol, however, may down-regulate the glucocorticoid receptor on immune cells, leading to glucocorticoid resistance and to sustained inflammation (Stark et al., 2001). Pro-inflammatory cytokines themselves also may induce glucocorticoid resistance (Miller et al., 2009a)
disrupting the social status of rodents leads to dysregulation of the inflammatory response via glucocorticoid resistance (Avitsur et al., 2001). Exposure to social-evaluative threat in humans also promotes glucocorticoid resistance (Dickerson et al., 2009)
rejection-related SAM activation could contribute to sustained inflammation through binding of catecholamines on immune cells and through sympathetic innervation of immune organs (Bierhaus et al., 2003; Elenkov et al., 2000b; however, see Elenkov et al., 2000a)
plausible pathways exist by which rejection may accelerate telomere shortening
Although biological reactivity to stress may be altered by exposure to prior stressors occurring at any time, early life stress may be particularly hazardous. Indeed, humans may experience the effects of environmental insults as early as gestational day 14, when in utero blood flow is established (Goodman and Brand, 2009). Fetuses of stressed mothers can be exposed to high levels of corticotrophin-releasing factor (CRF), potentiating reactivity to stress over the life course (Lupien et al., 2009)… Early maltreatment also predicts elevations in inflammation in adulthood (Danese et al., 2008)
individuals exposed to early parental loss, abuse, or neglect exhibit lower levels of pre-onset stress than those without such early adversity (Harkness et al., 2006)… Exposure to early interpersonal loss, therefore, may sensitize people uniquely to subsequent situations involving loss or rejection
Caspi et al. (2003) found that individuals with one or two copies of the short variant of 5-HTTLPR were more likely than long/long carriers to develop depression following early maltreatment or recent life stress
individuals with risk variants of the 5-HTTLPR polymorphism have been shown to exhibit greater amygdala responses to emotional stimuli (Munafò et al., 2008); greater amygdala reactivity, in turn, has been associated with greater chronicity of depression (Dannlowski et al., 2008). Short/short carriers also appear to exhibit potentiated biological responses to stress, as indexed by greater and more prolonged elevations in cortisol (Gotlib et al., 2008)
Smith MA, Davidson J, Ritchie J, Kudler H, Lipper S, Chappell P, Nemeroff C. 1989. “The corticotropin-releasing hormone test in patients with posttraumatic stress disorder” Biol Psychiatry 26(4):349-55
http://www.ncbi.nlm.nih.gov/pubmed/2548631
The PTSD patients had a significantly lower ACTH response to CRH compared to a control group of normal volunteers. Blunted ACTH responses occurred in patients with PTSD alone, as well as those PTSD patients who also had major depression
Smith ME. 2005. “Bilateral hippocampal volume reductions in adults with post-traumatic stress disorder: a meta-analysis of structural MRI studies” Hippocampus 15(6):798-807
http://www.ncbi.nlm.nih.gov/pubmed/15988763/
13 studies … 215 patients and 325 control… studies varied with respect to participant age, gender distribution, source of trauma, severity of symptoms, duration of disorder, the nature of the control groups, and the methods for volumetric quantification.
pooled effect size calculations indicated significant volume differences in both hemispheres. On average PTSD patients had a 6.9% smaller left hippocampal volume and a 6.6% smaller right
differences were smaller when comparing PTSD patients with control subjects exposed to similar levels of trauma, and larger when comparing PTSD patients to control subjects without significant trauma exposure
Smith G. 2014. “Estimating the population attributable fraction for schizophrenia when Toxiplasma gondii is assumed absent in human populations” Preventive Veterinary Medicine
http://www.sciencedirect.com/science/article/pii/S016758771400347X
the life-time mean population attributable fraction was estimated to be 21.4%, but it could not be ruled out that it could be as high as 30.6% or as low as 13.7%
Smith M, Cobia D, Reilly J, Gilman J, Roberts A, Alpert K, Wang L, Breiter H, Csernansky J. 2015. “Cannabis-related episodic memory deficits and hippocampal morphological differences in healthy individuals and schizophrenia subjects” Hippocampus DOI: 10.1002/hipo.22427
http://onlinelibrary.wiley.com/doi/10.1002/hipo.22427/full in dropbox
Authors include Jodi Gilman, lead author of “quite possibly the worst paper” of 2014 https://liorpachter.wordpress.com/2014/04/17/does-researching-casual-marijuana-use-cause-brain-abnormalities/
44 healthy controls (CON), 10 subjects with a CUD history (CON-CUD), 28 schizophrenia subjects with no history of substance use disorders (SCZ), and 15 schizophrenia subjects with a CUD history (SCZ-CUD)
Shape differences observed in CON-CUD were associated with poorer EM performance, while shape differences observed in SCZ-CUD were associated with a longer duration of CUD and shorter duration of CUD remission. A past history of CUD may be associated with notable differences in hippocampal morphology and EM impairments among adults with and without schizophrenia. Although the results may be compatible with a causal hypothesis, we must consider that the observed cannabis-related shape differences in the hippocampus could also be explained as biomarkers of a neurobiological susceptibility to poor memory or the effects of cannabis
The main effects of group and the group-by-hemisphere interaction were not significant for any comparison (P > 0.10; Table 2)
Although the main effect of group was nonsignificant (P = 0.16), the hippocampal volume was lower in SCZ-CUD compared with SCZ in the left (percent difference: −8.11%, d = −0.62), but not the right (percent difference: −4.58%, d = −0.31) hemisphere
we did not find differences in SCZ-CUD that were observed among CON-CUD
To conclude, the results suggest that a past CUD during adolescence may be associated with differences in hippocampal morphology and EM impairments in young adulthood even after a prolonged period of abstinence
Smith P, Homish G, Collins R, Giovino G, White H, Leonard K. 2014. “Couples’ marijuana use is inversely related to their intimate partner violence of the first 9 years of marriage” Psychol Addict Behav 28(3): 734-42
http://www.ncbi.nlm.nih.gov/pubmed/25134048
more frequent marijuana use by husbands and wives predicted less frequent IPV perpetration by husbands. Husbands’ marijuana use also predicted less frequent IPV perpetration by wives
There was a significant positive association between wives’ marijuana use and wives’ IPV perpetration, but only among wives who had already reported IPV perpetration during the year before marriage
overall inverse association between marijuana use and IPV perpetration in newly married couples
Smolen J, Maini R. 2006. “Interleukin-6: a new therapeutic target” Arthritis Res Ther 8(6): 407
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3226077/
IL-6 is expressed in a large proportion of cells in rheumatoid synovial tissue but not in osteoarthritis [6,7]. It has been shown that TNF levels are twice as high in RA as in psoriatic arthritis and 9–10 times higher in RA than in osteoarthritis [8]; this is also true of IL-6
TNF blockade reduces IL-6 serum levels [10], indicating an inter-relationship between these cytokines. It is important to note, however, that this is a downstream effect and that TNF blockade does not directly block the activity of IL-6
Blocking the IL-6 receptor with tocilizumab leads to significant improvement in signs and symptoms, and leads to reduction in and almost normalization of the acute-phase response
Sneider J, Gruber S, Rogowska J, Silveri M, Yurgelun-Todd D. 2013. “A preliminary study of functional brain activation among marijuana users during performance of a virtual water maze task” J Addict 461029
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3742334/
brain activation during the performance of a virtual analogue of the classic Morris water maze task in 10 chronic marijuana (MJ) users compared to 18 nonusing (NU) comparison subjects.
While task performance on learning trials was similar between groups, MJ users demonstrated a deficit in memory retrieval
NU subjects exhibited greater activation in the right parahippocampal gyrus and cingulate gyrus compared to the MJ group for the Retrieval-Motor Control contrast (NU > MJ)
To qualify for study entry, MJ smokers had to have smoked MJ a minimum of 2500 times, used MJ at least five of the last seven days prior to the study visit, test positive for urinary cannabinoids, and meet DSM-IV criteria for MJ abuse on the day of scanning. MJ users were asked to refrain from smoking for 12 hours
MJ users reported consuming 4.4 ± 4.3 alcoholic beverages per week, while NU participants reported consuming 1.8 ± 2.5 alcoholic beverages per week (F(1,26) = 4.2, P = .05). Four MJ users reported recent nicotine use (ranging from 1 pack per day; 1 pack every 2 weeks; 1 pack per month; occasional/social use). NU adults did not report any use of nicotine
All participants were free of Axis I diagnosis, except the MJ group, who were required to meet criteria for MJ abuse
No significant differences were observed between the groups for IQ, as measured by WASI, or for mood, as measured by the HAM-A, or on PANAS (positive or negative affect subscales) (). Further, no significant performance differences were detected between the MJ and NU groups on the Mental Rotation Test Total Score, or on the SBSOD Total Score
There was a significant group effect for latency to first movement. MJ users demonstrated shorter latencies to first movement (3.2 sec ± 1.2) relative to NU (4.6 ± 1.6) (F(1, 26) = 5.9, P < .05) and longer path lengths to reach the hidden platform (MJ: 1.7 ± 1.5 versus NU: 0.8 ± 0.4; F(1, 26) = 4.8, P < .05). There were no significant group differences for navigation latency (MJ users: 15.1 ± 12.9; NU: 9.9 ± 5.5).
There was a significant preference observed for the use of a direct versus a nondirect strategy during the Probe Trial in the NU group, with 78% (n = 14) employing a direct search strategy and 22% (n = 4) employing a nondirect strategy (χ2(1,18) = 5.6, P < .05). A strategy preference was not observed in the MJ group, 50% (n = 5) utilized a direct strategy and 50% (n = 5) utilized a nondirect strategy
NU, relative to MJ group, demonstrated greater BOLD activation in the bilateral inferior frontal pars triangularis and bilateral inferior frontal pars opercularis, left superior frontal gyrus, left superior frontal pars orbitalis, bilateral middle frontal gyrus, right pallidum, and right putamen for the Retrieval-Motor Control contrast (). However, the MJ group relative to NU did not show greater BOLD activation in any region
hyperactivation of brain areas suggest functional compensation and possible neural recruitment of additional brain areas
no significant group differences in BOLD activation were detected in the hippocampus
Brain activation changes associated with MJ use appear to be task specific, with some studies demonstrating hyperactivation, suggestive of increased recruitment, and other studies demonstrating hypoactivation, suggestive of inefficient neural networking [42, 51, 52]
MV: increased activation is a “functional compensation” whereas reduced activation is a “subtle deficit” even if neither are associated with impairment. Any effect, regardless of direction, is automatically interpreted as evidence of harm.
Snitz B, MacDonald A III, Carter C. 2006. “Cognitive deficits in unaffected first-degree relatives of schizophrenia patients: a meta-analytic review of putative endophenotypes” Schizophr Bull 32(1): 179-194
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2632195/
Cognitive deficits may index genetic liability for schizophrenia and are candidate endophenotypes for the illness
review reports mean effect sizes for 43 cognitive test scores from 58 studies of cognitive performance in the unaffected adult relatives of schizophrenia patients
largest effect sizes seen in complex versions of continuous performance tasks, auditory verbal learning, design copy tests, and category fluency
Three study design features were found to have significant effects on overall effect size magnitude: groups unmatched on education, groups unmatched on age, and asymmetric psychiatric exclusion criteria
moderate to large effect sizes in patient versus control differences in global and selective verbal memory, nonverbal memory, bilateral and unilateral motor performance, visual and auditory attention, general intelligence, spatial ability, executive function, language, and interhemispheric tactile transfer.6
findings indicate that relatives are also impaired, albeit to a lesser degree than patients, on a wide array of cognitive tasks (e.g., 8–14), although findings of no impairment have also been reported (e.g., 15–16)
The documented presence or absence of SPD or schizotypal symptoms also did not show an effect. There is much variability in the literature with regard to associations between cognitive deficits and schizotypy, a candidate endophenotype in its own right.41–42
Cognitive impairments in healthy relatives are clearly less severe than in their ill probands, which comes as little surprise. However, the range of ES magnitude for healthy relatives is comparable to that reported for cognitive deficits in some clinical populations, such as people occupationally exposed to mercury,49 symptomatic individuals infected with HIV who have not yet developed AIDS,50 and patients with mild head injury (overall d = 0.41 within 7 days of injury).51
Cognitive deficits, perhaps those involving executive control, working memory, and inhibition, in particular, may continue to prove valuable in the search for specific genes conferring risk for schizophrenia.
Solowij N, Michie P, Fox A. 1995. “Differential impairments of selective attention due to frequency and duration of cannabis use” Biol Psychiatry 37(10): 731-9
http://www.ncbi.nlm.nih.gov/pubmed/7640328
It has been suggested that the hippocampus plays a role in excluding extraneous stimuli during concentration of attention and that cannabinoids may disinhibit septal-hippocampal inputs to the reticular activating system, resulting in failure to habituate to irrelevant stimuli (Miller and Branconnier 1983)
ERP components are markers of specific stages of information processing, reflecting the nature, timing, and duration of cognitive processes (Hillyard and Kutas 1983; Naatanen 1990). In ERP studies of auditory selective attention, directing attention to a particular channel of information results in the onset of a negative shift of the ERP waveform, which is referred to as processing negativity (PN)Processing negativity is evident in the auditory system as early as 60-80 ms poststimulus, reflecting the selection of relevant from irrelevant sources of information. Ongoing negativity beyond 600 ms in the ERP waveform to attended stimuli is referred to as late PN and reflects the maintenance and rehearsal of the attentional trace
The other component of interest is the P300, a large positive peak which is elicited by infrequent stimuli in the attended channel when a response is required… the latency of the P300 component reflects the time taken to evaluate a stimulus, while its amplitude reflects the nature of stimulus evaluation processes
In a previous study (Solowij et al 1991), we compared ERPs recorded during a complex auditory selective attention task from a small and heterogenous group of long-term cannabis users and a nonuser control group. The results indicated that compared to controls, cannabis users showed larger processing negativity to an irrelevant source of information, reflecting an inability to filter out extraneous information at an early stage of processing, and reduced P300 amplitude, interpreted as a dysfunction in the allocation of attentional resources
regular use of at least once a month for 3 years. Detailed drug histories were taken; according to DSM-III-R definitions (American Psychiatric Association 1987), all subjects could be labeled “cannabis dependent” or “cannabis abusers.”
The cannabis user group (n = 32) had used cannabis for a mean of 6.69 years (range: 3 to 28 years) at a mean frequency of 12 days per month (range: once/month to daily use)
group was split at the median on both frequency (light: –<2/week vs. heavy: ->3/week) and duration (short: 3-4 years vs. long: ->5 years) of cannabis use.
cannabis users consumed more alcohol per month than controls
Controls had slightly more years of education than users [F(1,46) = 9.54; p < .005]; however, full-scale IQ, assessed by the NART (Nelson 1984), did not differ between groups based on duration of use relative to controls [F(2,45) = 2.04; p >. 1], nor between groups based on frequency of use [F(2,45) = 2.30; p >. 1 ]. A trend toward lower IQ overall in the cannabis users compared to controls [F(1,46) = 3.56; p > .06] indicated that the relationships between 1Q and ERP components needed to be examined in light of any group differences. The actual difference in IQ was very small, at 116.3 for users compared to 119.5 for controls
Subjects were excluded from the sample if they had a history of any psychiatric or neurological disorders, head injury, or significant use of any other drugs (-> once/month), or alcohol abuse. Cannabis users were instructed to abstain from cannabis and alcohol for 24 hours prior to testing
relationship between IQ and PN remained significant after controlling for the effects of age (r = .52; p < .05), whereas there was no relationship between age and PN after controlling for IQ (r = .10). PN and IQ were unrelated in the cannabis user group (r = -. 10)
cannabis users overall showed increased processing negativity to tones which matched the target on location but not pitch. When the cannabis user group was split according to frequency and duration of use, it became apparent that PN was greatly affected by duration of use
Long-duration users differed significantly from controls at frontal and central sites below a NART score of 42 (estimated full-scale IQ of 121), while short-duration users differed significantly from controls below a NART score of 35 (estimated full-scale IQ of 115) [F(1,28) = 4.20; p < .05 for both comparisons]. These results raise the possibility of more severe consequences from cannabis use for those of lower IQ However, the cut-off levels below which group differences were apparent are within the superior range of IQ for the long-duration user group and the high-average range for IQ for short-term users. This indicates that the effects of cannabis use are more apparent in users of low IQ when cannabis has only been used for a few years, but that when cannabis has been used for many years, its effects override any effect of IQ. This might also be interpreted to suggest that only users of superior IQ may be able to compensate and overcome subtle impairments. The positive relationship between PN and IQ in controls is puzzling. One interpretation of this relationship is that brighter subjects may be better able to process multiple sources of stimuli without any resultant loss of performance. It is important to note that the cannabis users tended to have a lower mean IQ than the controls but showed larger PN to pitch irrelevant tones. If the effects were solely due to IQ differences, one would have expected the reverse, i.e., cannabis users would have shown smaller PN to pitch-irrelevant tones than controls
contrary to our previous findings, there was no difference between cannabis users and controls in P300 amplitude…however, when the cannabis user group was split according to frequency and duration of use, it became apparent that P300 latency was greatly affected by frequency of use. The peak of the P300 component appeared to be delayed by over 100 ms in the heavy user group compared to both light users and controls
Group multiple comparisons at each site revealed that group differences were largest at the parietal site contralateral to the stimulated ear: heavy vs. light users: [F(1,25)= 17.49; p < .0005]; heavy users vs. controls: F(1,27) = 5.15; p < .05]. Light users did not differ from controls [F(1,26) = 2.25, p > .1]. The mean latency of the P300 peak across the three parietal scalp sites was 584 ms in heavy users and 498 ms each in light users and controls
no differences between groups based on duration of use [F(2,39) = i.20; p > .3] and no interactions between frequency and duration [F(1,23) = 1.93; p >. 1]
heavy-frequency use was found to prolong stimulus evaluation time, measured by P300 latency; long-duration use impaired the ability to effectively focus attention and reject irrelevant information, evidenced by increased processing negativity to irrelevant stimuli
it is the control group that differs in the two studies• The control group in the original study showed a much larger P300 than did controls of the current study (see Solowij et al 1991 Fig. 1, for comparison). No apparent differences in the characteristics of the two samples could explain this, indicating that effects of cannabis use on P300 amplitude are less robust than effects on PN. Similarly, P300 latency was not significantly delayed in the original study (there was a nonsignificant trend)
60% of the sample had no detectable cannabinoid metabolites in their urines
Neither time since last use, nor urinary cannabinoid metabolite levels, were related to the duration effect on processing negativity
Solowij N, Stephens R, Roffman R, Babor T, Kadden R, Miller M, Christiansen K, McRee B, Vendetti J. 2002. “Cognitive functioning of long-term heavy cannabis users seeking treatment” JAMA 287(9): 1123-1131
http://jama.jamanetwork.com/article.aspx?articleid=194703
Long-term cannabis users performed significantly less well than shorter-term users and controls on tests of memory and attention. On the Rey Auditory Verbal Learning Test, long-term users recalled significantly fewer words than either shorter-term users (P = .001) or controls (P = .005); there was no difference between shorter-term users and controls. Long-term users showed impaired learning (P = .007), retention (P = .003), and retrieval (P = .002) compared with controls. Both user groups performed poorly on a time estimation task (P<.001 vs controls). Performance measures often correlated significantly with the duration of cannabis use, being worse with increasing years of use, but were unrelated to withdrawal symptoms and persisted after controlling for recent cannabis use and other drug use
Scientific evidence from past research clearly showed that gross impairment related to chronic cannabis use did not occur but was inconclusive with regard to the presence of more specific deficits.5,6
compare (1) long-term users with a mean of 23.9 years of regular cannabis use; (2) shorter-term users with a mean of 10.2 years of regular use; and (3) nonusers of cannabis. Key confounding variables (age, IQ, other drug use) were controlled through matching or statistical methods
Cannabis users were included if they had used cannabis regularly for at least 3 years, were currently using at least once a week, were seeking treatment to assist them to cease or reduce their use of cannabis, and were willing to participate in the treatment program offered. Participants were excluded if they had ever had a serious illness or injury that may have affected the brain, any psychotic disorder, met a current DSM-IV diagnosis of dependence on any other drug or alcohol, or had a poor command of the English language
Overall, users and controls did not differ in age, but long-term users were significantly older and shorter-term users were significantly younger than controls (P<.001).
The mean estimated full-scale IQ (FSIQ) did not differ between the 3 groups based on duration of cannabis use. The majority of the sample (82.4% long-term, 88.2% shorter-term users) reported experiencing problems with memory, attention, or concentration, which they attributed to their use of cannabis.
Users first tried cannabis at a mean age of 15.3 (SD, 2.6) years with regular use (at least twice a month) commencing at age 17.5 (SD, 3.2) years. Cannabis had been used on a median 29 of the past 30 days (range, 1-30). Almost the entire sample (98%) met the DSM-IV criteria for cannabis dependence. The median amount of cannabis smoked per week was 1 quarter of an ounce (range, 0.01-2.00 oz) with 2 average-sized joints typically smoked per day (range, 0.12-20.00). None of these cannabis-use parameters differed between the long- and shorter-term user groups
Participants were required to abstain from cannabis for at least 12 hours prior to testing and to provide 2 urine samples (1 the night before testing, another during the test session). The median self-reported time since last use of cannabis was 17 hours (range, 7-240 hours)
At the time of testing, 70% of the sample reported that they were not experiencing any discomfort after abstaining from cannabis. Twice as many shorter-term users than long-term users (P = .03) reported mild withdrawal symptoms such as cravings, irritability, depression, anxiety, sleep, or appetite disturbances
Speed of Comprehension Cannabis user groups did not differ from controls in the number of items completed (range, 23-100) but users overall made more errors (P = .03) (range, 0-5). These results suggest that cannabis users are more likely to sacrifice accuracy for speed
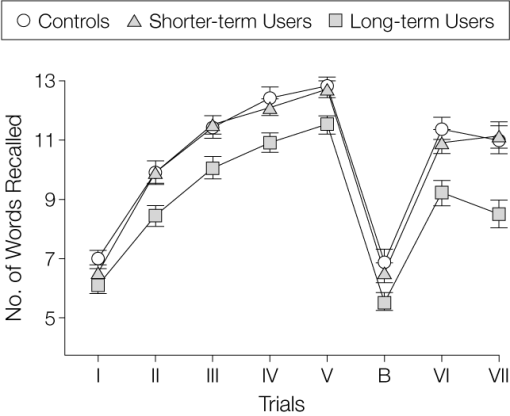
The proportion of subjects with a very poor learning ability (acquisition ❤ words over 5 trials) was greater among long-term users (13.7%) than controls (0%) (P = .007) but not shorter-term users (5.9%). The proportion of long-term users recalling fewer than 10 words on trial V (27.5%) was more than among shorter-term users (8.5%) or controls (3.0%) (P = .002). Significantly more long-term users (23.5%) lost 3 or more words over the 20-minute delay between trials VI and VII than shorter-term users (4.3%) or controls (3.0%) (P = .003). Long-term users showed a smaller primacy effect in the serial position curve than either other group (P = .02). Groups did not differ in the recency effect or in words recalled from the middle of the list.
Stroop: Cannabis users did not differ significantly from controls after inclusion of covariates in any condition or on interference scores
There were no significant group differences on any Wisconsin Card Sorting Test (WCST) measure but a trend on one: long-term users failed to maintain the set more often than shorter-term users (P = .05) or controls (P = .07). Research suggests that this measure best represents attentional dysfunction.39 There was no evidence of impaired performance with increasing years of cannabis use after controlling for covariates.
Groups did not differ in the time taken to complete any trial of the Alphabet Task or in the number of items correct in the Omitted Numbers task
Cannabis users differed from controls (P<.001) in Time Estimation Task A where they estimated the time taken to complete the preceding (Omitted Numbers) task. Both long- and shorter-term users underestimated the time by about one third of the actual time taken (64.4 seconds) and differed significantly from controls (P = .01 and P<.001, respectively). Groups did not differ in the simple and brief warned passive Time Estimation Task B or Time Production, where they could use strategies such as counting. Time estimation measures did not correlate with duration of cannabis use
Auditory Consonant Trigrams: Long-term users recalled significantly fewer items than shorter-term users (P = .007), controls (P = .002), and published norms48 on only the 9-second delay condition. The number of items recalled did not correlate with duration of cannabis use. In the general population, the greater the delay interval the worse the performance. In cannabis users, this general pattern was apparent, though there was greater interference at the shorter-delay interval than would be expected
Paced Auditory Serial Addition Test Long-term users had slower processing rates than shorter-term users on trial 1 (P = .007), with trends on trial 2 (P = .03) and the total processing rate across all trials (P = .02). Group differences on all other measures failed to reach significance but the performance of the long-term users was poorer in comparison with one set of norms49 but not another
When the sample was split at the median for time since last use or level of urinary cannabinoid metabolite on the day of testing and analyzed by ANCOVA, there were no differences on any measure between those who had used cannabis within the past 17 hours and those who had used cannabis 17 or more hours ago, or those with high vs low levels of urinary metabolites and no interactions with duration of cannabis use
Long-term users with a mean 24 years of regular cannabis use performed significantly less well on tests of memory and attention than nonuser controls and shorter-term users with a mean of 10 years’ use. The greatest impairment on almost every measure was from the RAVLT, indicating a generalized memory deficit with impaired learning, retention, and retrieval. Long-term users recalled 2.5 fewer words than controls on the delayed recall trial where 49% of the long-term users’ scores were more than 1 SD, and 21.6% were more than 2 SDs, below the control mean and normative data
These results do not indicate a severe memory problem
Despite this and a range of up to 17 years of cannabis use in the shorter-term user group, they differed significantly from controls only on time estimation.
our results suggest that shorter-term cannabis users are not impaired to an extent that would interfere with cognitive functioning in their daily lives
While most cannabis users cease using in their mid-20s to late 20s, approximately 20% continue to use through their 30s and beyond
a slightly higher proportion of shorter-term vs long-term users reported experiencing cognitive problems, yet shorter-term users mostly did not differ from controls on the cognitive tests. Nevertheless, it is possible that long-term cannabis users in the community who are not seeking treatment may not experience impairments to the same degree as those assessed in this study
Solowij N, Yucel M, Respondek C, Whittle S, Lindsay E, Pantelis C, Lubman D. 2011. “Cerebellar white-matter changes in cannabis users with and without schizophrenia” Psychological Medicine 41: 2349-2359
http://www.ncbi.nlm.nih.gov/pubmed/21466751
Seventeen patients with schizophrenia and 31 healthy controls were recruited ; 48 % of the healthy group and 47 % of the patients were long-term heavy cannabis users (mean 19.7 and 17.9 years near daily use respectively). Cerebellar measures were extracted from structural 3-T magnetic resonance imaging (MRI) scans using semi-automated methods, and examined using analysis of covariance (ANCOVA) and correlational analyses
Cerebellar white-matter volume was reduced in cannabis users with and without schizophrenia compared to healthy non-users, by 29.7 % and 23.9 % respectively, and by 17.7 % in patients without cannabis use. Healthy cannabis users did not differ in white-matter volume from either of the schizophrenia groups. There were no group differences in cerebellar grey matter or total volumes
Right-handed males… otherwise healthy long-term heavy cannabis users (THC ; n =15) ; healthy non-user controls (CON ; n=16) ; cannabis users with schizophrenia (SZ+THC ; n=8) and non-users with schizophrenia (SZ–THC ; n=9)… Cannabis users with and without schizophrenia had similar levels of current use and very extensive cannabis use histories (near daily use for 9–32 years)… alcohol use was limited to<24 standard drinks per week
All groups were matched on age (range 21–60 years), pre-morbid IQ (National Adult Reading Test) and years of edu-cation
All patients were stabilized on atypical antipsychotic medication (ex-cept for one currently antipsychotic free in the SZ–THC group). Estimated chlorpromazine equivalent doses did not differ between SZ+THC and SZ–THC (median 200 mg in each group ; p=0.54)
Hamilton Depression Rating Scale (HAMD) and Global Assessment of Functioning (GAF), also differed between the four groups (Table 1). HAMD scores were significantly higher in THC (t =2.60,p=0.013) and in SZ+THC ( t=2.33,p=0.024) compared to CON, and GAF scores were lower in both schizophrenia groups compared to both THC (SZ+THC : t =5.53,p<0.001 ; SZ–THC : t =5.21,p <0.001) and CON (SZ +THC : t =7.63,p<0.001 ; SZ–THC : t =7.39,p<0.001) and in THC compared to CON (t =2.45,p=0.018)
636 (565) cones/month. 19.7 (7.3) years
determined a significant overall difference between the four groups in white-matter volume [F(3, 38) =4.23,p=0.011], but groups did not differ in grey matter [ F (3, 38) =0.55,p=0.65] or total cerebellar volume [ F (3, 42)=1.68, p=0.19]. Whole-brain white-matter volume did not differ be-tween groups
GAF did not correlate with cer-ebellar volumetric measures (p>0.13), but the HAMD score correlated inversely with both white-matter ( p= 0.058) and total cerebellar volume (p=0.01), but not grey matter ( p=0.60) in the entire sample
The SAPS composite score correlated positively with white matter in the SZ+THC group (Spearman’s r=0.72,p=0.022) but inversely with total volume in the SZ–THC group (Spearman’s r=–0.70, p=0.018) No associations between SAPS composite scores and cerebellar volumes were evident in the healthy can-nabis users, and SANS composite scores did not correlate with cerebellar volumes in any group
In the SZ–THC group, there were also trends towards smaller total cerebellar volume ( r =0.62, p=0.078) and smaller grey-matter volume ( r =0.69,p=0.058) the earlier the age at onset of illness, with no such association in SZ +THC
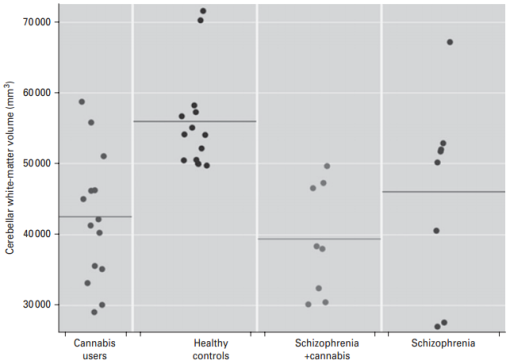
cannabis use may confer a relatively greater adverse effect on cerebellar white matter than schizophrenia… As a cross-sectional investigation, however, it cannot be determined whether these anomalies resulted from or preceded cannabis use
Because white-matter volume did not correlate with any specific measure of cannabis use, the findings cannot be interpreted as dose related. It remains possible that smaller cer-ebellar white-matter volume preceded cannabis use and may somehow be related to a propensity or pre-disposition to use cannabis.
Sonuga-Barke E, Castellanos F. 2007. “Spontaneous attentional fluctuations in impaired states and pathological conditions: a neurobiological hypothesis” Neuroscience and Behavioral Reviews 31(7): 977-86
http://adhdnet.com/wp/papers/Sonuga-Barke.pdf
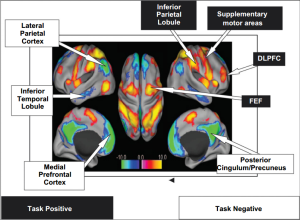
In traditional accounts, fluctuations in sustained and focused attention and associated attentional lapses during task performance are regarded as the result of failures of top-down and effortful higher order processes. The current paper reviews an alternative hypothesis: that spontaneous patterns of very low frequency (<0:1 Hz) coherence within a specific brain network (‘default-mode network’) thought to support a pattern of generalized task-non-specific cognition during rest, can persist or intrude into periods of active task-specific processing, producing periodic fluctuations in attention that compete with goal-directed activity
The capacity to effectively establish and actively maintain a sustained focus of mental activity during goaldirected tasks is both a necessary aspect of successful information processing (Van der Linden et al., 2005) and a key element of human productivity in post-industrial economies (Lanham, 2006). This is underlined by the fact that deficits in sustained/focused attention are an inherent feature of a number of debilitating psychiatric conditions (Brooks et al., 2006; Liu et al., 2002; Swaab-Barneveld et al., 2000)
This hypothesis predicts that disturbances in attention due to interference by the default mode would produce a distinctive pattern of slowly fluctuating attention with episodic lapses and a pattern of periodic and transitory performance deficits (e.g., manifest as increases in reaction time and increased frequency of errors)
The striking degree of anti-correlation of the taskpositive and task-negative networks (Fox et al., 2005; Fransson, 2005, 2006) suggests a reciprocal relationship and a potential antagonism in terms of the psychological functions these networks serve (Fox et al., 2005). The tight temporal linkage between task-positive and task-negative components implies that they can be considered as constituents of a single complex network
The task-negative elements of the network have been associated with an introspective attentional orientation related to mentalising and emotional processing (Gusnard et al., 2001) and the maintenance of a sense of self (Gusnard, 2005). These elements can be classified as ‘stimulus-independent thoughts’ and in as much as they are more or less incompatible with goal-directed activity, represent a potential source of cognitive task interference (cf. Smallwood and Schooler, 2006)
Mason et al. (2007) who demonstrated an association between stimulus-independent thoughts during task performance and both default-mode network activation and participant’s reports of the subjective experience of their ‘‘mind’s wandering’’
low frequency fluctuations synchronize activity within the taskpositive and task-negative networks implicated in defaultmode activity (Fransson, 2005, 2006)
extrospective task positive modes of attention are interspersed during periods of selfreflection and/or introspection to ensure a generalized periodic attentiveness to the possibility of unexpected and novel events in their external environment. An alternative point of view sees this toggling between introspective and extrospective modes of attention as the platform on which a constant updating of decision making frame of reference is based, as individuals weigh up of short and longer term outcomes within the environment
Given the traditional bias towards task-focused attention this antagonism is usually presented in terms of the way that episodes of task-independent thought in the form of self-referential introspection and reflection constrain the ability to focus and sustain attention during goal-directed tasks and so undermine task performance
effective attention is an emergent property which depends on selforganizing interactions (Van Orden et al., 2003) between intrinsic and spontaneous network activity. Deficiencies in self-organizing interactions would be one source of ineffective performance (Fox et al., 2005)
we have posited a threshold under which persisting task-negative low frequency oscillations will not impinge on task-related attention, but over which periodic attentional lapses will occur. An alternative hypothesis is that impact of task-negative activation during task performance operates in a continuous and linear way and that subtle interference in task performance can occur even with relatively small fluctuations in power of the default mode
will vary as a function of both state- and task-related factors (e.g., fatigue, boredom and levels of intrinsic motivation) as well as individual traits (e.g., levels of resting state coherence and state regulation deficits).
State-related variables such as those described above will inevitably interact with trait-type features in determining the liability to default-mode interference… Individuals differ in their tolerance for boredom and the extent to which they are intrinsically motivated by a particular task… ADHD represents an obvious place to start when considering the role of default-mode interference in individual differences of this sort (but see Kennedy et al., 2006; for a consideration in relation to autism)
Imaging studies, both structural and functional, provide some support for this model with evidence of reduced volumes in dorsolateral prefrontal cortex and the neostriatum (caudate/putamen; Castellanos et al., 2002; Shaw et al., 2006) and reduced activations during executive performance in the ventrolateral and dorsolateral prefrontal cortex (Rubia et al., 2005; Durston et al., 2003) as well as the neo-striatum (i.e., caudate and putamen; Vaidya et al., 2005)
only about half the children with ADHD displayed a deficit on any one executive task and very few subjects showed a pervasive pattern of EF deficits (Nigg et al., 2005). This suggests: (i) that while many children with ADHD show evidence of executive dysfunction on neuropsychological tasks, many do not (around 50% on any particular task; Willcutt et al., 2005)
ADHD is an umbrella construct, which while clinically useful, subsumes multiple groups of patients with distinctive aetiological and pathophysiological profiles
ADHD children’s performance would be more variable than that of controls and that this variability would have a low frequency time signature that is approximately synchronized with spontaneous lapses in attention to task, increases in stimulusindependent introspection and increased oscillatory power and coherence with the task-negative component of the default network in the low frequency signature (approximately between 0.01 and 0.1 Hz)
reaction times of children with ADHD are not only typically slower than controls but also almost always more variable (Klein et al., 2006).
Tian et al. (2006) who found significantly greater low frequency resting state connectivity for the ADHD group than in controls, in dorsal anterior cingulate cortex, thalamus, cerebellum, insula and brain stem. This finding is consistent with predictions of greater resting state inertia in ADHD
Sørensen H, Mortensen E, Schiffman J, Reinisch J, Maeda J, Mednick S. 2010 “Early developmental milestones and risk of schizophrenia. A 45-year follow-up of the Copenhagen Perinatal Cohort” Schizophr Res 118(0): 41-47
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3676634/
Five developmental milestones in particular (smiling, lifting head, sitting, crawling, and walking) differed significantly. Individuals who later developed psychiatric disorders other than schizophrenia reached most developmental milestones earlier than those who developed schizophrenia, but later than the controls. The two psychiatric groups only differed significantly with respect to age of walking without support
D’Souza D, Ranganathan M, Braley G, Gueorguieva R, Zimolo Z, Cooper T, Perry E, Krystal J. 2008. “Blunted psychotomimetic and amnestic effects of Δ-9-Tetrahydrocannabinol in frequent users of cannabis” Neuropsychopharmacology 33(10): 2505-2516
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3799954/
frequent users of cannabis are either inherently blunted in their response to, and/or develop tolerance to the psychotomimetic, perceptual altering, amnestic, endocrine and other effects of cannabinoids.
relative to controls, frequent users showed blunted responses to the psychotomimetic, perceptual altering, cognitive impairing, anxiogenic, and cortisol increasing effects of Δ-9-THC but not to its euphoric effects
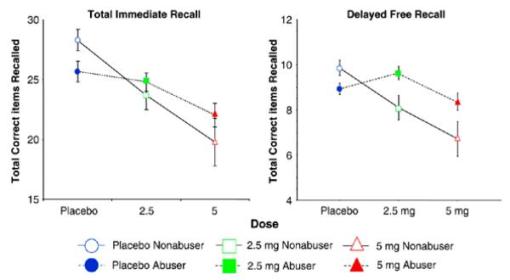
Δ-9-THC did not impair recognition recall
Distractability: Δ-9-THC increased omission errors only in controls (ATS = 7.86, df = 1, p = 0.01 for 5 mg vs. placebo comparison and ATS=10.41, p=0.002 for 5 mg vs. 2.5 mg comparison) without increasing commission errors.
Importantly however, despite having lower I.Q. scores and worse recall at baseline (placebo condition), frequent users had blunted Δ-9-THC induced immediate recall impairment relative to controls. Another intriguing finding of this study is that frequent users had betterdelayed recall under the influence of 2.5 mg Δ-9-THC, relative to the placebo condition
ndividuals at risk for nicotine and alcohol addiction have blunted sensitivity to the “negative” effects, but heightened sensitivity to the “positive” effects of these drugs (Conrod et al. 2001; Eissenberg & Balster 2000; Newlin & Thomson 1990; Pollock 1992; Schuckit 1985b; Schuckit 1994; Schuckit et al. 1991a;Schuckit et al. 1991b; Schuckit et al. 1996)
while frequent users showed blunted responses to the “negative” or ”undesirable” effects of Δ-9-THC, they were no different from controls in their response to Δ-9-THC induced “high”.
Sorrells S, Caso J, Munhoz C, Sapolsky R. 2009. “The stressed CNS: when glucocorticoids aggravate inflammation” Neuron 64(1): 33-9
http://www.sciencedirect.com/science/article/pii/S0896627309007430
Glucocorticoids (GCs) are hormones released during the stress response that are well known for their immunosuppressive and anti-inflammatory properties; however, recent advances have uncovered situations wherein they have effects in the opposite direction.
The canonical physiological stress response begins when the brain detects a homeostatic challenge and activates the sympathetic nervous system (SNS), which releases the catecholamines epinephrine (E) and norepinephrine (NE). This is followed by the slower activation of the hypothalamic-pituitary-adrenal (HPA)-axis: hypothalamic secretion of corticotropin-releasing hormone (CRH)
CORT secretion into the bloodstream peaks just prior to waking, with a 5-fold variation in levels across the circadian cycle. In response to substantial stressors, CORT secretion increases approximately an order of magnitude
we define “acute stress” as a stressor of a few hours. If such stress is repeated daily for several days we will refer to it as “subacute stress,” and if it persists for weeks to months, then it is termed “chronic stress.”
GCs, while renowned for their anti-inflammatory actions, are not uniformly anti-inflammatory, including in the context of CNS inflammation. Both GCs and stress (in a GC- and GR-dependent manner) can augment aspects of inflammation in the CNS, and these GC actions can exacerbate injury-induced neuronal death
Whether GCs increase or decrease CNS inflammation depends on the dose, timing, duration of GC exposure, and the type of GC, since these steroids are a heterogeneous group of compounds.
Spechler P, Orr C, Chaarani B, Kan K-J, Mackey S, Morton A, Snowe M, Hudson K, Althoff R, Higgins S, Cattrell A, Flor H, Nees F, Banaschewski T, Bokde A, Whelan R, Buchel C, Bromberg U, Conrod P, Frouin V, Papadopolous D, Gallinat J, Heinz A, Walter H, Ittermann B, Gowland P, Paus T, Poustka L, Martinot J-L. 2015. “Cannabis use in early adolescence: evidence of amygdala hypersensitivity to signals of threat” Developmental Cognitive Neuroscience 16: 63-70
http://www.sciencedirect.com/science/article/pii/S1878929315000857
we used an fMRI affective face processing task to compare a large group (n = 70) of 14-year olds with a history of cannabis use to a group (n = 70) of never-using controls matched on numerous characteristics including IQ, SES, alcohol and cigarette use. The task contained short movies displaying angry and neutral faces. Results indicated that cannabis users had greater reactivity in the bilateral amygdalae to angry faces than neutral faces, an effect that was not observed in their abstinent peers. In contrast, activity levels in the cannabis users in cortical areas including the right temporal-parietal junction and bilateral dorsolateral prefrontal cortex did not discriminate between the two face conditions, but did differ in controls
Controls were identified and matched on sex, handedness, age, verbal comprehension and perceptual reasoning IQ, pubertal development, socioeconomic status, and site… also matched on lifetime alcohol and cigarette use
With regard to the amygdala findings, it is unclear whether amygdala hypersensitivity preceded cannabis use or was a consequence of use since this was a cross-sectional study. If amygdala hypersensitivity preceded use, which might seem most plausible given the low levels of reported use, then it’s possible that these individuals may have been inclined to self-medicate for the drug’s acute anxiolytic effects (Phan et al., 2008). Consistent with this interpretation, recent evidence has identified altered angry face processing in the ventromedial PFC (vmPFC) to predict future binge drinking (Whelan et al., 2014) and the vmPFC is part of a brain circuit that attenuates amygdala activity (Banks et al., 2007 and Urry et al., 2006)
Greater signal change in the amygdala, specifically during affective face processing, is exhibited by children with anxiety (Thomas et al., 2001), and adults with major depressive disorder (Drevets, 2001, Fu et al., 2008 and Sheline et al., 2001) and generalized social phobia (Evans et al., 2008 and Phan et al., 2006)
we stress that we are unable to claim cannabis use caused amygdala hypersensitivity to negative affect. To investigate this question, longitudinal data analysis will inform whether hypersensitivity to threat signals precedes use or is a consequence of use
Squeglia L, Jacobus J, Brumback T, Meloy M, Tapert S. 2014. “White matter integrity in alcohol-naive youth with a family history of alcohol use disorders” Psychol Med 44(13): 2775-2786
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4134398/
youth with family history of alcoholism had higher fractional anisotropy and axial diffusivity, and lower radial and mean diffusivity, than FHN youth in 19 clusters spanning projection, association and interhemispheric white matter tracts
More mature neural features could be related to more precocious behaviors, such as substance use initiation
Neuropsychological studies have shown that alcohol-naive FHP children tend to perform worse on tasks of attention (Tarter et al. 1989; Ozkaragoz et al. 1997; Corral et al. 1999), visuospatial functioning (Ozkaragoz et al. 1997; Corral et al. 1999), memory (Ozkaragoz et al. 1997) and executive functioning (Giancola et al. 1993; Harden & Pihl, 1995; Corral et al. 2003) than FHN youth, with these cognitive deficits persisting into young adulthood (Lovallo et al. 2006; Acheson et al. 2011)
Gray matter maturation typically follows an inverted U-shaped pattern, with increases in gray matter until early adolescence (roughly peaking at age 11–13 years), followed by decreased volume into early adulthood (Huttenlocher, 1990; Sowell et al. 1999; Giedd, 2004; Gogtay et al. 2004; Giedd et al. 2009). In contrast, white matter continues to increase linearly, well into early adulthood (Giedd, 2004; Lenroot & Giedd, 2006; Bava et al. 2010; Yap et al. 2013). Increased white matter volume is believed to be related to progressive myelination of axons during adolescence, and is associated with greater connectivity between brain regions (Pfefferbaum et al. 1994, Sowell et al. 2004; Jernigan & Gamst, 2005; Hüppi & Dubois, 2006), smoother and more efficient communication between frontal–subcortical brain regions (Luna & Sweeney, 2004) and improved cognitive performance (Bava et al. 2010)
Alcohol-naive FHP youth have shown both higher FA (left posterior superior corona radiata/centrum semiovale) and lower FA (right anterior superior corona radiata, left superior longitudinal fasciculus, right extreme capsule, left inferior longitudinal fasciculus, left anterior limb of the internal capsule, and right optic radiation)
greater FA in frontal white matter tracts in at-risk adolescents when compared with controls, including youth who meet criteria for attention deficit/hyperactivity disorder (ADHD) (Li et al. 2010) and conduct disorder (Sarkar et al. 2013), as well as teens who engage in more dangerous and risky behaviors (Berns et al. 2009). Increases in white matter integrity observed during adolescence (Bava et al. 2010; Yap et al. 2013)
Alcohol consumption has been associated with lower white matter integrity in chronic alcoholic adults (Pfefferbaum et al. 2006a,b) and binge-drinking teens (McQueeny et al. 2009)
Despite alcohol-naive FHP youth showing greater white matter coherence, this advantage, or possible vulnerability, attenuates after youth initiate heavy substance use during adolescence. Specifically, previous studies have consistently found that heavy substance-using teens show decreasing FA compared with their non-using counterparts after alcohol and marijuana initiation (McQueeny et al. 2009; Bava et al. 2013;Jacobus et al. 2013a,b)
Stansfield K, Ruby K, Soares B, McGlothan J, Liu X, Guilarte T. 2015. “Early-life lead exposure recapitulates the selective loss of parvalbumin-positive GABAergic internerurons and subcortical dopamine system hyperactivity present in schizophrenia” Translational Psychiatry 5: e522
http://www.nature.com/tp/journal/v5/n3/full/tp2014147a.html
Environmental factors have been associated with psychiatric disorders and recent epidemiological studies suggest an association between prenatal lead (Pb2+) exposure and schizophrenia (SZ). Pb2+ is a potent antagonist of the N-methyl-d-aspartate receptor (NMDAR) and converging evidence indicates that NMDAR hypofunction has a key role in the pathophysiology of SZ. The glutamatergic hypothesis of SZ posits that NMDAR hypofunction results in the loss of parvalbumin (PV)-positive GABAergic interneurons (PVGI) in the brain. Loss of PVGI inhibitory control to pyramidal cells alters the excitatory drive to midbrain dopamine neurons increasing subcortical dopaminergic activity.
adolescence rats chronically exposed to Pb2+ from gestation through adolescence exhibit loss of PVGI in SZ-relevant brain regions. PV and glutamic acid decarboxylase 67 kDa (GAD67) protein were significantly decreased in Pb2+ exposed rats with no apparent change in calretinin or calbindin protein levels suggesting a selective effect on the PV phenotype of GABAergic interneurons. We also show that Pb2+ animals exhibit a heightened locomotor response to cocaine and express significantly higher levels of dopamine metabolites and D2-dopamine receptors relative to controls indicative of subcortical dopaminergic hyperactivity. Our results show that developmental Pb2+exposure reproduces specific neuropathology and functional dopamine system changes present in SZ. We propose that exposure to environmental toxins that produce NMDAR hypofunction during critical periods of brain development may contribute significantly to the etiology of mental disorders
Schizophrenia (SZ) is a devastating mental disorder believed to be the result of an adverse neurodevelopmental event with the clinical expression of the disease in adolescent and young adults
various aspects of the broad environment, such as nutritional deficiency, obstetric complications and influenza outbreaks, have been implicated in the etiology of SZ.3, 6, 7
Since the early 1970s, epidemiological studies originating from different parts of the world have consistently demonstrated detrimental associations of childhood lead (Pb2+) exposure on children’s intelligence, learning ability and neuropsychological development.8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23,24, 25 During the last decade, studies have provided evidence that childhood Pb2+intoxication is also associated with psychiatric disorders including major depression, anxiety, delinquency or conduct disorder, and SZ in adolescent and young adults who were exposed to Pb2+ as children.19, 20, 21, 22, 23, 24, 25
Human neuroimaging studies in Pb2+ exposed adolescent/young adults that have been followed since infancy support the epidemiological findings with evidence of gray matter and myelin changes in brain regions affected in mental disorders.26, 27, 28, 29, 30, 31 In particular, the medial prefrontal cortex (mPFC) and hippocampus (HIPP) appear to be the two brain regions that are consistently affected in both Pb2+ exposed children and in SZ.26, 27, 28, 29
extensive evidence that Pb2+ is a potent and selective antagonist of the NMDAR37, 38, 39, 40, 41 and chronic exposure to Pb2+ in early life arrests the developmental NR2B-to-NR2A NMDAR subunit switch in the rat brain.40,41 This is important because maturation and refinement of many brain circuits is coincident with the NMDAR subunit switch.42, 43, 44, 45
medial ganglionic eminence-derived parvalbumin (PV)-positive GABAergic interneurons (PVGI) that undergo NR2B-to-NR2A subunit switch with a functional change in their firing rate from slow to fast signaling properties.45In general, dysregulation of this important developmental NMDAR subunit switch is associated with neurological and mental disorders
One of the most robust and consistent findings documented in postmortem brain tissue from SZ subjects and in NMDAR antagonist animal models of SZ is the loss of PVGI in the mPFC and HIPP.48, 49, 50, 51, 52, 53 PVGI are fast-spiking GABAergic inhibitory neurons that synchronize pyramidal cell firing in the mPFC and HIPP, giving rise to gamma oscillations that are critical for cognitive function.54, 55Alterations in the gamma oscillations in SZ contribute significantly to the cognitive impairments characteristic of the disease.56, 57 Consistent with this notion, genetic and pharmacological studies in experimental animals support an important role of the NMDAR in PVGI function. For example, genetic ablation of the NR1 subunit of the NMDAR in forebrain PVGI58 or the administration of NMDAR antagonists59, 60,61, 62, 63 results in the loss of PV and glutamic acid decarboxylase (GAD67), markers for GABAergic interneurons in the rodent brain
hyperactivity of subcortical DAergic neurons is a consistent finding in drug-naive SZ patients measured by PET imaging.65, 66, 67, 68
Animal studies have shown that excess dopamine (DA) in the striatum can be induced by abnormal activity of pyramidal cells in the mPFC and HIPP by reducing their GABAergic inhibitory control.69, 70Therefore, the loss or dysfunction of inhibitory PVGI control to principal neurons in the mPFC and HIPP would be expected to result in disinhibition of pyramidal cells, the loss of their synchronized firing, altering the excitatory drive to midbrain dopamine neurons increasing subcortical DAergic activity.34
Pb2+ exposure paradigm does not produce an overt toxic effect on the body weight as we did not find a significant effect of the chronic Pb2+ exposure of the body weight of rats at PN50
Pb2+ exposed rats body weight was 313±11 g (mean±s.e.m.; n=30; P=0.2). At PN50, the resulting blood Pb2+ levels in these animals was 22.2±0.9 μg dl−1 (n=47) for Pb2+ exposed rats and 0.6±0.1 μg dl−1 (n=64) for control rats
We found significantly higher concentrations of the DA metabolites DOPAC (208% increase; P=0.0009) and HVA (146% increase; P=0.016) in the striatum of Pb2+ exposed rats relative to controls (Figure 5b), whereas DA (P=0.089), DOPAC/DA (P=0.099) and HVA/DA (P=0.099) ratio did not reach statistical significance after adjusting for multiple comparisons (Figure 5b). Our results also revealed that Pb2+ animals had an overall higher level of [3H]-raclopride-specific binding in the striatum (averaged dorsal and ventral striatum) than control rats (P=0.0242; Figure 5d). The mean±s.e.m. of [3H]-raclopride-specific binding in the striatum of control animals was 53.1±1.9 fmol mg−1 tissue (n=7) and 58.7±0.4 fmol mg−1 tissue (n=6) for Pb2+ exposed animals. This change represents a 10.5% increase in the level of total striatal D2R
animals chronically exposed to environmentally relevant levels of Pb2+during brain development express three key features that are observed in SZ: (1) the loss of PVGI in brain regions relevant to SZ pathology, (2) subcortical DAergic hyperactivity and (3) increased levels of D2R in the striatum. The current findings combined with previous studies showing impairments of hippocampal long-term potentiation,46 adult neurogenesis in the HIPP,94 prepulse inhibition of the startle response95 and cognitive function46, 47, 96 in similarly Pb2+ exposed and age-matched animals are consistent with what has been observed in the SZ brain and in NMDAR antagonist animal models of SZ
the Pb2+exposure paradigm and the resulting levels of Pb2+ in the blood of adolescent rats used in the present study are relevant to the blood Pb2+ levels that have been measured for decades in a significant percentage of children not only in the United States (http://www.cdc.gov/nceh/lead/data/StateConfirmedByYear1997-2001.htm), but also worldwide.41, 97 Consistent with this notion, recent estimates by the World Health Organization indicate that Pb2+ exposure accounts for 0.6% of the global burden of disease and contributes to ~600 000 new cases of children with intellectual disabilities every year (http://www.who.int/ipcs/lead_campaign/en/). Further, data from the National Health and Nutrition Examination Survey (NHANES) indicate that ~1% of women of childbearing age (15–44 years) in the United Sates have blood Pb2+ levels that are equal or greater than the current Center for Disease Control (CDC) level of concern (http://www.cdc.gov/nceh/lead/publications/leadandpregnancy2010.pdf). Therefore, a significant percentage of children worldwide continue to be at risk for neurodevelopmental problems resulting from Pb2+ exposure during critical periods of brain development and these effects may contribute significantly to mental disorders across the lifespan.
Stark J, Avitsur R, Padgett D, Campbell K, Beck F, Sheridan J. 2001. “Social stress induces glucocorticoid resistance in macrophages” American Journal of Physiology 280(6): R1799-R1805
http://ajpregu.physiology.org/content/280/6/R1799
Stress-induced levels of plasma glucocorticoid hormones are known to modulate leukocyte function
a psychosocial stressor induced glucocorticoid resistance in mouse splenic macrophages
Steffens M, Engler C, Zentner J, Feuerstein T. 2004. “Cannabinoid CB1 receptor-mediated modulation of evoked dopamine release and of anenylyl cyclase activity in the human neocortex” Br J Pharmacol 141(7)
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1574880/
Electrically evoked [3H]-DA release from human neocortical slices was inhibited by CP55.940 (IC50 6.76 nm, Imax 65%) and strongly enhanced by the cannabinoid receptor antagonist AM251. However, [3H]-DA release was not influenced in rat neocortex
K+-evoked [3H]-DA release in the presence of tetrodotoxin (TTX) was strongly inhibited by CP55.940 in humans, but not in rats.
CP55.940 inhibited forskolin-stimulated cAMP accumulation (IC50 20.89 nm,Imax 35%). AM251 blocked this effect and per se increased forskolin-stimulated cAMP accumulation by ∼20%.
Stein M, Koverola C, Hanna C, Torchia M, McClarty B. 1997. “Hippocampal volume in women victimized by childhood sexual abuse” Psychol Med 27(4):951-9
http://www.ncbi.nlm.nih.gov/pubmed/9234472/
Women who reported sexual victimization in childhood had significantly reduced (5% smaller) left-sided hippocampal volume compared to the non-victimized women. Hippocampal volume was also smaller on the right side, but this failed to reach statistical significance. Left-sided hippocampal volume correlated highly (rs = -0.73) with dissociative symptom severity, but not with indices of explicit memory functioning
Stein-Behrens B, Lin W, Sapolsky R. 1994. “Physiological elevations of glucocorticoids potentiate glutamate accumulation in the hippocampus” J Neurochem 63(2): 596-602
http://www.ncbi.nlm.nih.gov/pubmed/7913489/
Glucocorticoids (GCs) are secreted during stress and can damage the hippocampus over the course of aging and impair the capacity of hippocampal neurons to survive excitotoxic insults
GCs augment the extracellular accumulation of glutamate and aspartate in the hippocampus following kainic acid-induced seizures
incremental increases in GC concentrations cause incremental increases in glutamate accumulation before the kainic acid insult, as well as in the magnitude of the glutamate response to kainic acid. Elevating GC concentrations from the circadian trough to peak doubled cumulative glutamate accumulation, whereas a rise into the stress range caused a fourfold increase in accumulation
stress levels of GCs augmented glutamate accumulation even in the absence of an excitotoxic insult, perhaps explaining how sustained stress itself damages the hippocampus
Steiner M, Wotjak C. 2008. “Role of the endocannabinoid system in regulation of the hypothalamic-pituitary-adrenocortical axis” Progress in Brain Research vol 170: 397-432
http://www.ncbi.nlm.nih.gov/pubmed/18655899 in dropbox
CB1 binding leads to opening of inwardly rectifying potassium (Kir) channels and closing of N- or P/Q-type Ca2+ channels. Also coupling to mitogen-activated protein kinase (MAPK) and akt thymona viral oncogene (Akt) signalling, as well as to the
phosphoinositol pathway
CB1 receptor-mediated reduction of glutamate release from glutamatergic synapses, which innervate and activate parvocellular neurons of the PVN, is held responsible for glucocorticoid-mediated fast-feed-back regulation of the HPA axis
CB1 receptor mRNA is co-localized with CRH mRNA in more than 50% of CRH positive neurons in the hypothalamic nuclei of the PVN also the amygdala, prefrontal cortex and BNST … involved in stress integration ( Cota et al., 2007)
CB1 receptors have also be en shown to co-localize with CRH type 1 receptors (CRH -R1) in neurons, which express low levels of CB1 and which are likely of glutamatergic nature ( Hermann and Lutz, 2005)
CB1 receptors are expressed primarily presynaptically, whereas CRH-R1s are thought to be localized postsynaptically ( Chen et a l., 2004)
Whereas CB1 receptors in humans are localized in somatotrophs, lactotrophs, corticotropes and folliculostellate cells ( Pagotto et al., 2001), CB1 receptors in the rat and frog pituitary are localized mainly in gonadotrophs and lactotrophs and in frogs to some extent in thyrotrophs, but CB1 receptors are largely absent from corticotropes in both rats and frogs (Wenger et al., 1999; Cesa et al., 2002 ).
costimulation of CB1 receptors by a CB1 agonist had an additive effect on CRH-stimulated ACTH secretion, suggesting that CB1 receptor signalling in the human pituitary may actually aid in regulating ACTH release
cannabinoid-mediated inhibition of adrenaline secretion from isolated rat adrenal glands despite lack of CB1 expression, suggested that this effect is likely mediated via CB1 receptor-mediated inhibition of acetylcholine release from preganglionic sympathetic nerve terminals ( Niederho ffer et al., 2001)
ACTH secretion due to THC in rats. Inverse U curve of glucocorticoid response from THC
Whereas a low dose (0.03 mg/kg CP-55,940 ) decreased stress-induced corticosterone secretion, a higher dose (0.3 mg/kg ) increased stress-induced corticosterone secretion
PVN of the hypothalamus, the BNST and the central nucleus of the amygdala (CeA). Interestingly, also the paraventricular nucleus of the thalamus (De Fonseca et al., 1997 ; Patel et al., 1998; Arnold et al., 2001), and its projection areas, the nucleus accumbens shell (De Fonseca et al., 1997; Arnold et al., 2001) and the prefrontal cortex (Arnold et al., 2001) were strongly activated
the PVN receives both direct glutamatergic and GABAergic afferences from surrounding hypothalamic areas as well as indirect innervations from limbic brain regions with at least one trans-synaptic relay (for review see Herman et al., 2004)
cannabinoid agonism downregulates vassopressin and oxytocin (Di et al., 2005); oxytocin stimulates cannabinoid release (Hirasawa et al., 2004)
Other than for the restraining of glutamate release, glucocorticoid- or OXT-controlled endocannabinoid signalling at GABAergic synapses is expected to result in increased rather than decreased secretion of the neuropeptide from axon terminals. Moreover, endocannabinoids seem to mediate the inhibitory effects of a-melanocyte-stimulating hormone (a-MSH) on spontaneous activity of oxytocinergic neurons ( Sabatier and Leng, 2006 ; not shown), which may serve as a switch between dendritic and axonal oxytocin release (Sabatier et al., 2003 ). Taken into account that magnocellular neurons may contribute to HPA axis regulation (Engelmann et al., 2004), with differential contribution of dendritic and axonal neuropeptide release (Wotjak et al., 2002)
hypothalamoneurohypophyseal system
TRPV1 agonist capsaicin caused an increase in excitatory glutamatergic transmission within the PVN (Li et al., 2004)
female CB1 receptor knockout mice increase in basal corticosterone secretion at the peak but not the nadir of the circadian rhythm of corticosterone secretion (Cota et al., 2007 )
male, but not female, CB1 receptor knockout mice showed elevated basal corticosterone levels also towards the end of the dark phase
CB1 mutant mice on the CD1 background (Ledent et al., 1999 ), which showed either decreased (Uriguen et al., 2004), increased ( Haller et al., 2004) or similar (Aso et al., 20 08) corticosterone and/or AC TH levels as compared to wild-type mice (compare also Table 3). No basal changes of corticosterone levels have so far been detected in CB1 receptor mutant mice on the C57BL/6J background (Zimm er et al., 1999 ; Fride et al., 2005; Wade et al., 2006). To explain the dependency of endocannabinoid action on the circadian rhythm we propose that the concentration of corticosterone has to exceed a certain threshold at level of the PVN, before it may activate local endocannabinoid synthesis and release
increasing the firing activity of serotonergic neurons within the dorsal raphe nucleus (Gob bi et al., 2005; Bam bico et al., 2007)
activation of CB1 receptors within the dentate gyrus of the dorsal hippocampus promote active stress coping behaviour
amplification of endocannabinoid signalling at their site of action by blocking endocannabinoid uptake and degradation ameliorated the deleterious consequences of chronic stress
Stellwagen D, Beattie E, Seo J, Malenka R. 2005. “Differential regulation of AMPA receptor and GABA receptor trafficking by tumor necrosis factor α” Journal of Neuroscience 25(12): 3219-3228
http://www.jneurosci.org/content/25/12/3219.full
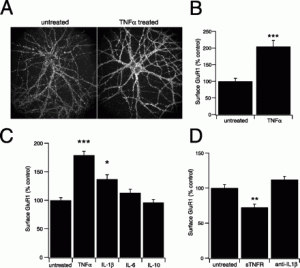
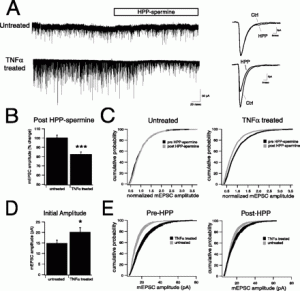
immune molecules such as cytokines are also thought to importantly contribute to the brain damage induced by a wide variety of neuronal insults as well as neurodegenerative diseases (New et al., 1998; Lock et al., 1999;Shohami et al., 1999; Nagatsu et al., 2000; Perry et al., 2001; Szelenyi, 2001), while also having some neuroprotective functions in certain circumstances (Morganti-Kossmann et al., 2002; Stoll et al., 2002)
tumor necrosis factor-α (TNFα) was found to be a novel, glia-released factor that increases the surface expression of AMPARs (Beattie et al., 2002
the TNFα-induced increase in the surface expression of AMPARs may contribute significantly to its putative role in mediating the brain damage resulting from a variety of pathological insults (Gelbard et al., 1993; Chao and Hu, 1994; Le et al., 1997; Hermann et al., 2001)
the increase in surface expression of AMPARs is attributable to the direct action of TNFα on neurons and not through another cell type
activation of TNFR1 is necessary for the maintenance of surface expression of AMPARs and is sufficient for their rapid exocytosis
although TNFα induced a robust twofold increase in GluR1 surface expression, in the same cells, it caused no significant increase in GluR2 surface expression
These results suggest that many, if not most, of the new surface AMPARs lack the GluR2 subunit, resulting in receptors that are calcium permeable, inwardly rectifying, and inhibited by polyamines such as HPP-spermine (Washburn et al., 1997;Mainen et al., 1998; Dingledine et al., 1999)
TNFα does not cause a change in the synaptic localization of NMDA receptors (Beattie et al., 2002)
there was no increase in the level of surface expression of GABAA receptors after 15 min of TNFα treatment compared with untreated cultures, despite a nearly twofold increase in the surface AMPARs that were measured in the same cells (Fig. 6A). In fact, TNFα caused a small but significant reduction in GABAA receptor surface expression (88 ± 4%; n = 237; p < 0.01) endocytosis
Treatment with TNFα nearly doubled the amount of internalized GABAAreceptors compared with untreated control cells (196 ± 17%; n = 55)
Modest increases in mEPSC amplitude combined with small decreases in mIPSC amplitude could combine to result in much larger changes in the ratio of excitatory to inhibitory synaptic transmission and thereby significantly affect network properties… The ratio of EPSP to IPSP amplitude from acute slices incubated in TNFα was increased more than twofold
TNFα is a proinflammatory cytokine that has been implicated in playing an important role in the neuronal damage caused by a variety of brain insults, such as stroke and head trauma, as well as that which occurs during neurodegenerative disorders (New et al., 1998; Lock et al., 1999; Shohami et al., 1999; Nagatsu et al., 2000; Perry et al., 2001; Szelenyi, 2001)
he findings presented here, specifically an increase in surface AMPARs, a change in their stoichiometry to make them Ca2+ permeable, and a decrease in inhibition, all will enhance the excitotoxicity that accompanies neuronal insults and some autoimmune and neurodegenerative diseases that may involve TNFα (Epstein and Gelbard, 1999; Weiss and Sensi, 2000)
Stinson F, Ruan W, Pickering R, Grant B. 2006. “Cannabis use disorders in the USA: prevalence, corelates and co-morbidity” Psychol Med 36(10: 1447-60
http://www.ncbi.nlm.nih.gov/pubmed/16854249/
The prevalence of 12-month and lifetime DSM-IV cannabis abuse (1.1% and 7.2%) exceeded the corresponding rates of cannabis dependence (0.3% and 1.3%). Being male, Native American, widowed/separated/divorced, and residing in the West increased the odds whereas being Black, Asian or Hispanic decreased the odds of cannabis abuse and dependence. Cannabis dependence was significantly associated with low income… Co-morbidity was high between cannabis use disorders and other Axis I and II disorders.
82% with lifetime marijuana use disorder (abuse or dependence) also met criteria for an alcohol use disorder, and 48% met criteria for nicotine dependence
Stochl J, Khandaker G, Lewis G, Perez J, Goodyer I, Zammit S, Sullivan S, Croudace T, Jones P. 2015. “Mood, anxiety and psychotic phenomena measure a common psychopathological factor” Psychol Med 45(7): 1483-93
http://www.ncbi.nlm.nih.gov/pubmed/25394403?dopt=Abstract
Psychotic phenomena are common in the general population but are excluded from diagnostic criteria for mild to moderate depression and anxiety despite their co-occurrence and shared risk factors
6617 participants aged 13 years from the Avon Longitudinal Study of Parents and Children (ALSPAC) birth cohort and 977 participants aged 18 years from the ROOTS schools-based sample
In both cohorts depression, anxiety and psychotic items were best represented as a bi-factor model with a single, unitary CMD factor on which psychotic items conveyed information about the more severe end (model 4)
Psychotic phenomena co-occur with depression and anxiety in teenagers and may be a marker of severity in a single, unitary dimension of CMD. Psychotic phenomena should be routinely included in epidemiological assessments of psychiatric morbidity, otherwise the most severe symptomatology remains unmeasured
Stokes P, Mehta M, Curran H, Breen G, Grasby P. 2009. “Can recreational doses of THC produce significant dopamine release in the human striatum?” Neuroimage 48(1): 186-90
http://www.ncbi.nlm.nih.gov/pubmed/19539765
“no”
Storr M, Yuce B, Andrews C, Sharkey K. 2008. “The role of the endocannabinoid system in the pathophysiology and treatment of irritable bowel syndrome” Neurogastroenterol Motil 20(8): 857-68
http://www.ncbi.nlm.nih.gov/pubmed/18710476
The endocannabinoid system is involved in the regulation of numerous gastrointestinal functions including motility, sensation and secretion under both physiological and pathophysiological conditions. Activation of cannabinoid (CB)(1) and CB(2) receptors under various circumstances reduces motility, limits secretion and decreases hypersensitivity in the gut. Drugs that alter the levels of endocannabinoids in the gut also reduce motility and attenuate inflammation
Straus M, Paschall M. 2009. “Corporal Punishment by Mothers and Development of Children’s Cognitive Ability: A Longitudinal Study of Two Nationally Representative Age Cohorts” Journal of Aggression, Maltreatment & Trauma 18:459-483
http://www.tandfonline.com/doi/abs/10.1080/10926770903035168 in dropbox
children 2-4 not hit gain 5.5 IQ relative to those hit 3 or more times per week (the norm in that age range); children 5-9 gained an average of almost 2 points. For 2-4, cognitive stimulation the largest factor, punishment second largest; for 5-9, punishment had the largest effect
18% of mothers hit the child during the interview. 93% in 2-4, 58% in 5-9, used CP. Average for those who used was 3.6 times per week. This rate is 10 times greater than mean number of times reported in prior 1-year recall study by same author
“selective inattention” (Dexter, 1958) allows us to continue to support CP because it avoids the necessity of facing up to the fact that almost all children are hit, and many frequently.
Strohbeck-Kuehner P, Skopp G, Mattern R. 2008. “Cannabis improves symptoms of ADHD” Cannabinoids 3(1):1-3
http://cannabis-med.org/data/pdf/en_2008_01_1.pdf
A 28-year old male, who showed improper behaviour and appeared to be very maladjusted and inattentive while sober, appeared to be completely inconspicuous while having a very high blood plasma level of delta-9- tetrahydrocannabinol (THC)… THC can have atypical effects and can even lead to an enhanced driving related performance
Sullivan J. 2000. “Cellular and molecular mechanisms underlying learning and memory impairments produced by cannabinoids” Learn Mem 7: 132-139
http://learnmem.cshlp.org/content/7/3/132.long
vitro experiments find that cannabinoid receptor activation reduces neurotransmitter release below the levels required to trigger long-term changes in synaptic strength in the hippocampus. Cannabinoids reduce glutamate release through a G-protein-mediated inhibition of the calcium channels responsible for neurotransmitter release from hippocampal neurons
Sullivan N, Elzinga S, Raber J. 2013. “Determination of pesticide residues in cannabis smoke” J Toxicol 2013: 378168
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3666265/
Recoveries of residues were as high as 69.5% depending on the device used and the component investigated, suggesting that the potential of pesticide and chemical residue exposures to cannabis users is substantial and may pose a significant toxicological threat in the absence of adequate regulatory frameworks
Surguladze S, Young A, Senior C, Brebion G, Travis M, Phillips M. 2004. “Recognition accuracy and response bias to happy and sad facial expressions in patients with major depression” Neuropsychology 18(2): 212-8
http://www.ncbi.nlm.nih.gov/pubmed/15099143?dopt=Abstract
depressed patients demonstrated subtle impairments in discrimination accuracy and a predominant bias away from the identification as happy of mildly happy expressions
Swartz J, Knodt A, Radtke S, Hariri A. 2015. “A neural biomarker of psychological vulnerability to future life stress” Neuron 85(3): 505-511
http://www.cell.com/neuron/abstract/S0896-6273(14)01164-7
Amygdala reactivity interacts with stress to predict internalizing symptoms
Amygdala reactivity predicted symptoms as much as 1 to 4 years after scanning
von Sydow K, Lieb R, Pfister H, Hofler M, Wittchen H. 2002. “What predicts incident use of cannabis and progression to abuse and dependence? A 4-year prospective examination of risk factors in a community sample of adolescents and young adults.” Drug Alcohol Depend 68(1):49-64
http://www.ncbi.nlm.nih.gov/pubmed/12167552
Incident predicted mainly by availability of drugs, peers’ drug use, a more ‘positive’ attitude towards future drug use, and regular previous use of licit drugs, while cannabis dependence was predicted primarily by parental death before age 15, deprived socio-economic status, and baseline use of other illicit drugs. unsatisfactory relationships with mothers a factor
Tait R, Mackinnon A, Christensen H. 2011. “Cannabis use and cognitive function: 8-year trajectory in a young adult cohort” Addiction 106: 2195-2203
http://www.ncbi.nlm.nih.gov/pubmed/21749524
We assessed cognitive performance with the California Verbal Learning Test (CVLT) (immediate and delayed), Spot-the-Word test (STW), Symbol Digit Modality test (SDMT) and Digit Backwards (DB)
‘never’ (n = 420) ‘remain light’ ( n = 71), ‘former light’ (n = 231), ‘remain heavy’ (n = 60), ‘former heavy’ ( n = 60) and ‘always former’ (since start of study) ( n = 657)
heavy users (at least weekly)
At baseline there were significant differences between cannabis groups on CVLT (immediate and delayed) and SDMT. However, after controlling for education, gender, gender ¥ group and gender ¥ wave, there were no significant between-group differences and only CVLT immediate recall reached adjusted statis-tically significant longitudinal change associated with changed cannabis use (group ¥ waveP = 0.007). Specifically, former heavy users improved their performance relative to remaining heavy users (estimated marginal means: former heavy 6.1–7.5: remain heavy 6.4–6.6)
a smaller proportion of females than males remaining heavy users of cannabis and remaining heavy users having the fewest years of education
On all measures the significance of cannabis group declined after controlling for the other variables, most notably on the SDMT — Symbol Digit Modalities Test
Only with respect to the immediate recall measure was there evidence of an improved performance associated with sustained abstinence from cannabis, with outcomes similar to those who had never used cannabis at the end-point. On the remaining cognitive measures, after con-trolling for education and other characteristics, there were no significant differences associated with cannabis consumption
Stereotype threat results from negative self-evaluation about group membership, with male cannabis users incurring greater decrements than female cannabis users, at least when the stereotypes are actively invoked [33]
adverse impacts of cannabis use on cognitive functions either appear to be related to pre-existing factors or are reversible in this community cohort even after potentially extended periods of use. These findings may be useful in motivating individuals to lower cannabis use, even after an extensive history of heavy intake
Takahashi R, Pamplona F, Fernandes M. 2005. “The cannabinoid antagonist SR141716A facilitates memory acquisition and consolidation in the mouse elevated T-maze” Neurosci Lett 380(3):270-5
http://www.ncbi.nlm.nih.gov/pubmed/15862900
memory improvement, observed when the drug was administered before (effect on memory acquisition/consolidation) or immediately after the training (effect on memory consolidation), but not before the test (effect on memory retrieval). Also, SR administration reversed scopolamine-induced amnesia
Tan H-T, Nicodemus K, Chen Q, Li Z, Brooke J, Honea R, Kolachana B, Straub R, Meyer-Lindenberg A, Sei Y, Mattay V, Callicott J, Weinberger D. 2008. “Genetic variation in AKT1 is linked to dopamine-associated prefrontal cortical structure and function in humans” J Clin Invest 118(6): 2200-2208
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2391279/
Cognitive performance linked to frontostriatal circuitry, prefrontal physiology during executive function, and frontostriatal gray-matter volume on MRI were altered in subjects with the AKT1 variation
significant epistasis with a functional polymorphism (Val158Met) in catechol-O-methyltransferase [COMT], a gene that indexes cortical synaptic dopamine
We studied AKT1 effects on human subjects at several phenotypic levels — 319 individuals in terms of cognitive tests, a subset of 32 individuals in terms of AKT1 protein expression in lymphoblasts, independent subsets of 46 and 68 individuals in terms of fMRI of brain, and an overlapping set of 171 individuals for structural brain MRI
Factor 4, comprising measures of IQ, executive function, and processing speed from subtests of the Wechsler Adult Intelligence Scale — Revised (WAIS-R), the Trail Making Test, and the Verbal and Category Fluency Test (33), was associated with significant AKT1 haplotypic variation, the strongest comprising SNPs rs3730358, rs1130233 (formerly aliased rs2498799 in the earlier literature), and rs2494732 (global P < 10–6, uncorrected). The specific GAC haplotype was associated with reduced factor 4 scores (P < 0.002, uncorrected, or P < 0.02, Bonferroni corrected for the 8 possible haplotype combinations in these 3 SNPs). Of the individual SNPs in this haplotype, a linear regression analysis revealed that the rs1130233 A allele was associated with reduced factor 4 scores… The neuropsychological measures comprising this cognitive factor reflect IQ, processing speed, and executive function engaging brain networks that include frontal cortex and striatum (36–39).
minor allele of SNP rs1130233, associated previously with reduced performance on cognitive tests and reduced AKT1 protein levels, engaged increased prefrontal activation, which suggests a relatively inefficient activation phenotype (Brodmann area [BA] 9: Montreal Neurological Institute coordinates –38, 30, 36 mm, z score (z) = 3.47; P = 0.0003, uncorrected, or P < 0.05, small-volume corrected according to the principles of Gaussian random fields [ref. 46]; Figure . This phenotype… has been linked to reduced cortical DA function in normal subjects (26, 28, 30, 47) and in patients with Parkinson disease (48, 49).
the AKT1 SNP rs1130233 minor allele was associated with relatively reduced gray-matter volumes in the bilateral caudate (coordinates –10, 7, 3 mm, z = 3.79, P < 0.05, small-volume corrected; coordinates 12, 7, 4 mm, z = 3.37, P < 0.05, small-volume corrected) and right prefrontal cortex (BA 47: 37, 18, –12 mm, z = 3.73, P < 0.05, small-volume corrected; Figure and Supplemental Table 6)
AKT1 genetic variation did not map strongly onto other cognitive factors engaging episodic memory or attention nor factors potentially representing other aspects of executive function (e.g., factors 2 and 5)
AKT-knockout mice had working memory behavioral deficits and evidenced parallel changes in prefrontal pyramidal dendritic ultrastructure and downstream alterations in the expression of genes controlling synaptic function, neuronal development, myelination, and actin polymerization in prefrontal cortex (7)
s1130233 has previously been implicated in AKT1 expression levels and in mediating radiation-induced apoptosis of lymphocytes (42). We confirmed its effect on the expression of AKT1 protein in lymphoblasts from a subgroup of the normal subjects in our sample and confirmed the directionality of the allelic association. Consistent with basic models suggesting deficits in AKT phosphorylation and signaling underlying cognitive deficits (5, 7, 8), the A allele at rs1130233 associated with poorer cognitive function and relative deficits in neuroimaging measures correspondingly evidenced reduced levels of AKT1 in the lymphoblasts of a subset of the same individuals
Chronic opiate exposure also has been shown to downregulate AKT signaling and lead to decreased DA cell size in ventral tegmental area (51)
our results support notions that genetic mechanisms modestly associated with clinical psychiatric syndromes are more powerfully examined through effects at the level of fundamental brain mechanisms and cognitive processes
Of note, modulating the AKT-related pathways in the direction of that apparently observed in psychosis is a major therapeutic goal for cancer (72). This not only adds to the interesting epidemiological observation that risk for schizophrenia and cancer might be inversely related (73) but should also encourage further preclinical and human studies specifying these pathways, implicated also in neurodegeneration (74)
Tao Y, Tang J, Chen Q, Guo J, Li L, Yang L, Feng H, Zhu G, Chen Z. 2015. “Cannabinoid CB2 receptor stimulation attenuates brain edema and neurological deficits in a germinal matrix hemorrhage rat model” Brain Res 1602:127-35
http://www.ncbi.nlm.nih.gov/pubmed/25625355
Germinal matrix hemorrhage (GMH) is one of the most common and devastating cerebrovascular events that affect premature infants, resulting in a significant socioeconomic burden
Up to 1h post-injury, the administration of JWH133 (1mg/kg, intraperitoneal injection) significantly attenuated brain edema at 24h post-GMH, which was reversed by a selective CB2R antagonist, SR144528 (3mg/kg, intraperitoneal injection). Long-term brain morphology and neurofunctional outcomes were also improved. In contrast, JWH133 did not have a noticeable effect on the hematoma volume during the acute phase. These data also showed that microglia activation and inflammatory cytokine (TNF-α) release were significantly inhibited by JWH133 after GMH. This current study suggests a potential clinical utility for CB2R agonists as a potential therapy to reduce neurological injury and improve patient outcomes after GMH
Tashkin DP. 2013. “Effects of Marijuana Smoking on the Lung” Annals of the American Thoracic Society 10(3):239-247
habitual use of marijuana alone does not appear to lead to significant abnormalities in lung function when assessed either cross-sectionally or longitudinally, except for possible increases in lung volumes and modest increases in airway resistance of unclear clinical significance
the accumulated weight of evidence implies far lower risks for pulmonary complications of even regular heavy use of marijuana compared with the grave pulmonary consequences of tobacco.
Teufel C, Subramaniam N, Dobler V, Perez J, Finnemann J, Mehta P, Goodyer I, Fletcher P. 2015. “Shift toward prior knowledge confers a perceptual advantage in early psychosis and psychosis-prone healthy individuals” PNAS 112(43): 13401-13406
http://www.pnas.org/content/112/43/13401.full
Current models of brain function view perception as a combination of two distinct sources of information: bottom-up sensory input and top-down influences from prior knowledge… we identified unmedicated individuals who experience early psychotic symptoms but fall below the threshold for a categorical diagnosis… there was a shift in information processing favoring prior knowledge over incoming sensory evidence
the degree of psychosis proneness in healthy individuals, and, specifically, the presence of subtle perceptual alterations, is also associated with stronger reliance on prior knowledge. Although, in the current experimental studies, this shift conferred a performance benefit, under most natural viewing situations, it may provoke anomalous perceptual experiences
vision seems to play a prominent role in the development of psychosis given that basic visual symptoms identified before illness onset are one of the most powerful predictors of the emergence of later psychotic disorders (14)
Schizotypy refers to a personality measure that has established predictive value for psychotic and other mental illnesses (18). Although it has been traditionally considered a specific risk measure for schizophrenia, more recently it has been proposed to reflect a general psychosis proneness
growing evidence suggesting that psychosis lies on a continuum with normality (19, 20) and is associated with a range of different psychiatric disorders (15, 21). According to this perspective, existing diagnostic categories group biologically heterogeneous syndromes with potentially different pathophysiological mechanisms into one disorder (22); this may obfuscate our attempts to understand the neurobiological underpinnings of mental illness
Such a result is rare and revealing in that it highlights a specific information-processing atypicality rather than a general performance deficit
it has been hypothesized that a single core disturbance relating to the balance between bottom-up and top-down processing can explain both the hallucinatory experiences and the bizarre delusional beliefs of psychotic patients (8, 11)
supports the idea that the putative atypicality underlying the emergence of perceptual psychotic experiences relates directly to normal function of the system. In other words, the potential for psychotic experiences such as hallucinations might be a logical consequence of the way in which our brain deals with the inherent ambiguity of sensory information by incorporating prior knowledge into our perceptual processing… within a hierarchical and recurrent information-processing system such as the human brain, an imbalance at any level will, in time, propagate up and down the hierarchy and affects the whole system (8, 30), a notion that might ultimately account for atypicalities in both lower-level perceptual processing and higher-level belief formation in severe mental illnesses and psychosis proneness (30)
he integration of bottom-up and top-down signals is mediated by the relative amount of information each of these components provides: the stronger the sensory evidence is relative to prior knowledge, the more it will impact on the final processing output; conversely, if prior knowledge provides a relatively greater amount of information it will be weighted more strongly
could either be due to atypically strong knowledge representations or, alternatively, to an unusually noisy sensory system. Given well-established psychotic deficits in early visual processing as discussed below, the latter explanation might be more likely
Some authors argue that, on a neuronal level, the bottom-up/top-down balance is implemented by neuromodulatory gain control of feedforward and feedback neuronal circuitries; adopting this approach, psychosis may thus be understood in terms of atypicalities in these control mechanisms (35). Although some models have focused on the emergence of delusions as a consequence of enhanced gain of feed-forward connections (and thus of the bottom-up signal) (8, 35, 36), the current findings suggest that a shift in the opposite direction—i.e., either enhanced gain of feedback connections or reduced gain of feedforward connections—may account for perceptual changes associated with psychosis. Given that our study did not directly measure neuromodulatory gain control, this assertion is speculative
atypicalities within the visual system result in the outputs of the perceptual analysis being less well structured and less adaptive. This could lead to an increased reliance on sources of information from outside the visual system to structure visual percepts and might make psychotic patients “hungry” for prior object knowledge. It is this latter effect that was demonstrated in the current study.
Tims FM, Dennis ML, Hamilton N, Buchan BJ, Diamond G, Funk R, Brantley LB. 2002. “Characteristics and problems of 600 adolescent cannabis users in outpatient treatment.” Addiction 97(s1): 46-57
http://onlinelibrary.wiley.com/doi/10.1046/j.1360-0443.97.s01.7.x/abstract?
46% met the DSM-IV criteria for substance dependence, while 50% met criteria for a diagnosis of abuse. Only 20% of the participants perceived any need for help with problems associated with their drug or alcohol use. Clients participating in the study typically presented multiple problems at treatment entry, most often including conduct disorder, attention deficit hyperactivity disorder (ADHD), internal (mental) distress, and physical health distress. The co-occurrence of conduct disorder and ADHD was found in 30% of the sample. Clients meeting criteria for substance dependence tended to have more co-occurring problems and significantly less denial at admission.
Tomoda A, Suzuki H, Rabi K, Sheu Y-S, Polcari A, Teicher M. 2009. “Reduced prefrontal cortical gray matter volume in young adults exposed to harsh corporal punishment” Neuroimage 47(Suppl 2): T66-T71
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2896871/
GMV was reduced by 19.1% in the right medial frontal gyrus (medial prefrontal cortex; MPFC, BA10) (P= 0.037, corrected cluster level), by 14.5% in the left medial frontal gyrus (dorsolateral prefrontal cortex; DLPFC, BA 9) (P = 0.015, uncorrected cluster level) and by 16.9% in the right anterior cingulate gyrus (BA 24) (P < 0.001, uncorrected cluster level) of HCP subjects. There were significant correlations between GMV in these identified regions and performance IQ on the WAIS-III. 10 point gap in verbal IQ
Traynelis S, Wollmuth L, McBain C, Menniti F, Vance K, Ogden K, Hansen K, Yuan H, Myers S, Dingledine R. 2010. “Glutamate receptor ion channels: structure, regulation and function” Pharmacol Rev 62(3): 405-496
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2964903/
Tzavara E, Wade M, Nomikos G. 2003. “Biphasic effects of cannabinoids on acetylcholine release in the hippocampus: site and mechanism of action” J Neurosci 23(28):9374-84
http://www.ncbi.nlm.nih.gov/pubmed/14561865
biphasic, dose-dependent, effect on hippocampal ACh: a low (0.5 mg/kg, i.p.) and a high (5 mg/kg, i.p) dose of WIN55,212-2 induces a transient stimulation and a prolonged inhibition of hippocampal ACh efflux, respectively.
Inhibition can be blocked by intrahippocampal rimonabant or pertussis toxin; stimulation can be blocked by the same in septum, in which the majority of cholinergic cell bodies projecting to the hippocampus reside
stimulatory and inhibitory effects of WIN55,212-2 on hippocampal ACh involve dopamine D1 and D2 receptor activation, respectively, given that pretreatment with D1 and D2 receptor antagonists prevents the respective actions of WIN55,212-2
in vivo biphasic effects of CB1R agonists on hippocampal ACh release result from a differential, functional association of anatomicaly distinct subpopulations of CB1-Gi coupled receptors to neurotransmitter systems that have opposing effects on ACh release
Tzilos G, Cintron C, Wood J, Simpson N, Young A, Pope H Jr, Yurgelun-Todd D. 2005. “Lack of hippocampal volume change in long-term heavy cannabis users” Am J Addict 14(1):64-72
http://www.ncbi.nlm.nih.gov/pubmed/15804878/
smokers displayed no significant adjusted differences in volumes of gray matter, white matter, cerebrospinal fluid, or left and right hippocampus. hippocampal volume in cannabis users was not associated with age of onset of use not total lifetime episodes of use. These findings are consistent with recent literature suggesting that cannabis use is not associated with structural changes within the brain as a whole or the hippocampus in particular.
Uhlhaas P, Singer W. 2010. “Abnormal neural oscillations and synchrony in schizophrenia” Nat Rev Neurosci 11(2): 100-13
http://www.ncbi.nlm.nih.gov/pubmed/20087360 in dropbox
Converging evidence from electrophysiological, physiological and anatomical studies suggests that abnormalities in the synchronized oscillatory activity of neurons may have a central role in the pathophysiology of schizophrenia
positive symptoms may be associated with circumscribed increases in oscillatory activity that may reflect the read-out of stored experiences
Uhlhaas P. 2012. “Dysconnectivity, large-scale networks and neuronal dynamics in schizophrenia” Curr Opin Neurobiol 23(2): 283-90
http://www.ncbi.nlm.nih.gov/pubmed/23228430 in dropbox
Ujike H, Morita Y. 2004. “New perspective sin the studies on endocannabinoid and cannabis: cannabinoid receptors and schizophrenia” J Pharmacol Sci 96(4): 376-81
http://www.ncbi.nlm.nih.gov/pubmed/15613777
CB1 density elevated in prefrontal cortex in schizophrenia; anandamide elevated in CSF. CNR1 gene 9-repeate allele AAT-repeat polymorphism predicts 2.3-fold higher susceptibility to schizophrenia.
Varese F, Smeets F, Drukker M, Lieverse R, Lataster T, Viechtbauer W, Read J, van Os J, Bentall R. 2012. “Childhood Adversities Increase the Risk of Psychosis: A Meta-analysis of Patient Control, Prospective-and Cross-sectional Cohort Studies” Schizophrenia Bulletin 38(4): 661-71
http://schizophreniabulletin.oxfordjournals.org/content/early/2012/03/28/schbul.sbs050.full
18 case-control studies (n = 2048 psychotic patients and 1856 nonpsychiatric controls), 10 prospective and quasi-prospective studies (n= 41 803) and 8 population-based cross-sectional studies (n = 35 546). There were significant associations between adversity and psychosis across all research designs, with an overall effect of OR = 2.78 The estimated population attributable risk was 33% (16%–47%)
Vatner S, Pachon R, Vatner D. 2015. “Inhibition of adenylyl cyclase type 5 increases longevity and healthful aging through oxidative stress protection” Oxidative Medicine and Cellular Longevity article ID 250310
http://www.hindawi.com/journals/omcl/2015/250310/
Mice with disruption of adenylyl cyclase type 5 (AC5 knockout, KO) live a third longer than littermates. The mechanism, in part, involves the MEK/ERK pathway, which in turn is related to protection against oxidative stress. The AC5 KO model also protects against diabetes, obesity, and the cardiomyopathy induced by aging, diabetes, and cardiac stress and also demonstrates improved exercise capacity. All of these salutary features are also mediated, in part, by oxidative stress protection
inhibition of AC5’s action to prolong longevity and enhance healthful aging, as well as its mechanism through resistance to oxidative stress, is unique among all of the nine AC isoforms
AC5 KO heart was protected against the stresses of chronic pressure overload [1] and chronic catecholamine stimulation [2, 3]
here is considerable need for a novel therapy that might mimic the phenotype of caloric restriction, without the negative aspects of limiting caloric intake. One such model is inhibition of AC5, which has a similar phenotype to caloric restriction [6], but the mice actually eat more than their wild type controls, while weighing less
Mitochondrial dysfunction and oxidative stress. During the aging process, ROS accumulation and DNA mutation in mitochondria induce oxidative stress, which results in the mitochondrial permeability transition pore (PTP) opening and electron transport chain deficiency, thereby leading to cell dysfunction and cell death. Inhibition of AC5 activates SIRT1/FoxO3a and Raf/MEK/ERK pathways, and both pathways upregulate the antioxidant, MnSOD, resulting in resistance to oxidative stress during aging
Oxidative stress is enhanced during the aging process; mitochondria produce less energy (ATP) and also increase the production of reactive oxygen species (ROS) as a product of aerobic metabolism (Figure 1). Furthermore the activity of free-radical scavenging enzymes changes with aging. One mechanism by which oxidative stress is increased in the aging tissue is through increased ROS induced apoptosis and necrosis by opening of the mitochondrial membrane permeability transition pore and release of apoptotic inducing factors, for example, cytochrome c [23]
considerable support for the regulation of oxidative stress by the Raf/MEK/ERK pathway [25]
final common pathway for premature cell death has been reported to be apoptosis, necrosis, and control of the cell cycle [26, 27] (Figure1), specifically an increase in the percentage of cells stopped at the G0/G1 phase of the cell cycle [26].
Based on the statistics of the CDC in 2010, the leading causes of death in humans in order of frequency are heart disease, cancer, chronic lower respiratory diseases, stroke, accidents, Alzheimer’s disease, diabetes, influenza and pneumonia, nephritis, nephrotic syndrome, and nephrosis. It is important to recognize that most of these diseases are related to oxidative stress. Such is the case for heart disease [67], cancer [68], COPD [69], stroke [70], Alzheimer’s [71], diabetes [72], and chronic kidney disease [73]
mechanisms by which obesity produces oxidative stress. First, adipose tissue produces certain bioactive substances called adipokines such as IL-6 and leptin, which induce the production of ROS. A second mechanism is that mitochondrial and peroxisomal oxidation of fatty acids can produce ROS in oxidation reactions. And third, there is an overconsumption of oxygen in the mitochondrial respiratory chain [85]… a study in humans confirmed the increased oxidative stress state seen in patients with obesity and diabetes, measuring urinary creatinine-8-epi-PGF2α as a marker of systemic oxidative stress [87]
AC/cAMP/PKA pathway negatively regulates the MEK-ERK signaling pathway [101]. The MEK/ERK signaling pathway is also one of the main stress signaling pathways and central mediators activated in response to oxidative damage [102, 103]
9 major mammalian isoforms of AC and many share regulation of the same tissue or organ in the body [126, 127]; yet they regulate in radically different ways. One example relates to the two major isoforms of AC in the heart, AC5 and AC6. Inhibiting AC6 shares none of the salutary features of inhibiting AC5
It is even more interesting and pertinent to the topic of oxidative stress that so little is known about AC isoforms and regulation of oxidative stress. Almost none of the other 8 AC isoforms have been shown to regulate oxidative stress (Table 2), with the tangential exception that AC1 can affect glutamate induced toxicity, which is related to oxidative stress, in cortical neurons. Although little is known about regulation of oxidative stress by the other AC isoforms, the reverse has been shown, that is, oxidative stress regulation of AC isoforms [135, 136]. In addition, the major mechanisms mediating oxidative stress in the AC5 KO, the SIRT1/FoxO3/MnSOD pathway, have also not been observed with the other AC isoforms (Table 2). It could well be that other AC isoforms do regulate oxidative stress but that this just has not been studied as of yet and would be an important future direction for AC research
Veen N, Selten J, van der Tweel I, Feller W, Hoek H, Kahn R. 2004. “Cannabis use and age at onset of schizophrenia” Am J Psychiatry 161(3): 501-6
http://www.ncbi.nlm.nih.gov/pubmed/14992976
Male patients were significantly younger than female patients at first social and/or occupational dysfunction, first psychotic episode, and first negative symptoms. Cannabis-using patients were significantly younger at these milestones than were patients who did not use cannabis. cannabis use, but not gender, made an independent contribution to the prediction of age at first psychotic episode: male cannabis users were a mean of 6.9 years younger at illness onset than male nonusers. age at first social and/or occupational dysfunction and the risk of developing negative symptoms before the first contact with a physician for treatment of possible psychotic disorder were predicted by gender, but not by cannabis use
Veldhuis W, van der Stelt M, Wadman M, van Zedelhoff G, Maccarrone M, Fezza F, Veldink G, Vliegenthart J, Bar P, Nicolay K, Di Marzo V. 2003. “Neuroprotection by the endogenous cannabinoid anandamide and arvanil against in vivo excitotoxicity in the rat: role of vanilloid receptors and lipxygenases” Journal of Neuroscience 23(1): 4127-4133
http://www.jneurosci.org/content/23/10/4127.long
pretreatment with the reduced 12-lipoxygenase metabolite of AEA, 12-HAEA, attenuated cytotoxic edema formation in a CB1 receptor-independent manner in the acute phase after intracranial injection of the Na+/K+-ATPase inhibitor ouabain; (2) the reduced 15-lipoxygenase metabolite, 15-HAEA, enhanced the neuroprotective effect of AEA in the acute phase; (3) modulation of VR1, as tested using arvanil, the VR1 agonist capsaicin, and the antagonist capsazepine, leads to neuroprotective effects in this model, and arvanil is a potent neuroprotectant, acting at both CB1 and VR1; and (4) the in vivo neuroprotective effects of AEA are mediated by CB1 but not by lipoxygenase metabolites or VR1
Verdoux H, van Os J. 2002. “Psychotic symptoms in non-clinical populations and the continuum of psychosis” Schizophr Res 54(1-2): 59-65
http://www.ncbi.nlm.nih.gov/pubmed/11853979
A growing body of evidence suggests that delusional or hallucinatory experiences are much more frequent in subjects from the general population than the prevalence of cases of psychotic disorders, thereby suggesting the existence of a symptomatic continuum between subjects from the general population and clinical cases of psychosis
Villarreal G, Hamilton D, Graham D, Driscoll I, Qualls C, Petropoulos H, Brooks W. 2004. “Reduced area of the corpus callosum in posttraumatic stress disorder” Psychiatry Res 131(3): 227-35
http://www.ncbi.nlm.nih.gov/pubmed/15465292S
12 adult patients with PTSD and 10 matched controls… the total CC area, absolute and normalized to total brain parenchyma, was smaller compared with control values… anterior mid-body (area 4) smaller in PTSD No differences were found in the rostrum (region 1), rostral body (region 3) or splenium (region 7)… not attributable to generalized white matter atrophy
Volkow N, Wolf A, Brodie J, Cancro R, Overall J, Rhoades H, Van Gelder P. 1988. “Brain interactions in chronic schizophrenics under resting and activation conditions” Schizophr Res 1(1): 47-53
http://www.ncbi.nlm.nih.gov/pubmed/3154506
Vyas A, Mitra R, Rao B, Chattarji S. 2002. “Chronic stress induces contrasting patterns of dendritic remodeling in hippocampal and amygdaloid neurons” The Journal of Neuroscience 22(15): 6810-6818
http://www.jneurosci.org/content/22/15/6810.long
chronic immobilization stress (CIS) induced dendritic atrophy and debranching in CA3 pyramidal neurons of the hippocampus. In striking contrast, pyramidal and stellate neurons in the basolateral complex of the amygdala exhibited enhanced dendritic arborization in response to the same CIS. Chronic unpredictable stress (CUS), however, had little effect on CA3 pyramidal neurons and induced atrophy only in BLA bipolar neurons
Vythilingam M, Heim C, Newport J, Miller A, Anderson E, Bronen R, Brummer M, Staib L, Vermetten E, Charney D, Nemeroff C, Bremner J. 2002. “Childhood trauma associated with smaller hippocampal volume in women with major depression” Am J Psychiatry 159(12):2072-80
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3230324/
restricting inclusion to women with a history of severe, recurrent, and prolonged abuse that occurred before puberty
depressed subjects with childhood abuse had an 18% smaller mean left hippocampal volume than the nonabused depressed subjects and a 15% smaller mean left hippocampal volume than the healthy subjects. Right hippocampal volume was similar across the three groups. The right and left hippocampal volumes in the depressed women without abuse were similar to those in the healthy subjects
unreported history of childhood abuse in depressed subjects could in part explain the inconsistencies in hippocampal volume findings in prior studies in major depressive disorder
826,000 cases of child maltreatment were reported in the United States in 1999 (30). Half of these cases were instances of neglect, 21% were instances of physical abuse, and 11% were instances of sexual abuse
40%–54% of the variance in hippocampal volume is genetically determined (66, 67)
Vythilingam M, Lawley M, Collin C, Bonne O, Agarwal R, Hadd K, Charney D, Grillon C. 2006. “Hydrocortisone impairs hippocampal-dependent trace eyeblink conditioning in Post-Traumatic Stress Disorder” Neuropsychopharmacology 31: 182-188
http://www.nature.com/npp/journal/v31/n1/full/1300843a.html
12 patients with PTSD and 12 age- and sex-matched healthy controls participated in a trace eyeblink test 6 h following intravenous administration of 30 mg of hydrocortisone. Spontaneous blink rates were similar between PTSD patients and healthy controls. There was no significant difference in the mean conditioned response between PTSD subjects and healthy controls under placebo conditions. Following hydrocortisone administration, only the PTSD patients demonstrated a significant reduction in conditioned response in contrast to healthy subjects who did not demonstrate any change. Patients with PTSD had increased glucocorticoid sensitivity in the focal brain regions mediating trace eyeblink conditioning
One of the advantages of evaluating eyeblink conditioning in neuropsychiatric populations is that it requires little cooperation from the subjects. Relative to other learning and memory tasks, results are less likely to be affected by nonspecific factors such as motivation.
Preclinical and emerging literature in humans suggest that trace eyeblink conditioning is dependent on both the cerebellum and the stress-sensitive hippocampal neurons in the CA1 and CA3 regions (Berger and Thompson, 1978; Clark and Squire, 1998; McEchron and Disterhoft, 1997; Moyeret al, 1990; Weiss et al, 1996)
Glucocorticoids play a critical role in modulating trace eyeblink conditioning since removal of both the adrenal cortex and medulla but not medulla alone prevents enhancement of trace eyeblink conditioning in male rats (Beylin et al, 2001; Beylin and Shors, 2003). Basal levels of glucocorticoids are also positively correlated with trace eyeblink conditioning (Wood et al, 2001)
Subjects with hypercortisolemia due to Cushing’s syndrome had fewer conditioned responses using a trace eyeblink conditioning paradigm compared to healthy subjects (Grillon et al, 2004)
The only study that evaluated eyeblink conditioning in PTSD used a delayed paradigm and reported impairments in both combat veterans with and without PTSD (Ayers et al, 2003). The results from this study are difficult to interpret because the experimental condition (paired CS-air puff) elicited less conditioned response compared to the control condition (unpaired CS-air puff) (Ayers et al, 2003), when the opposite should be expected
Trace eyeblink conditioning can also be pharmacologically manipulated similar to tasks evaluating learning and memory and is sensitive to the effects of glucocorticoids (Beylin and Shors, 2003)
mean conditioned response on placebo was similar in patients with PTSD and healthy controls. Hydrocortisone administration decreased the mean number of conditioned responses and the number of adaptive conditioned responses in PTSD but not in healthy controls, suggesting increased glucocorticoid sensitivity only in PTSD patients
preclinical studies support the role for CA3 and CA1 hippocampal neurons in mediating eyeblink trace conditioning (Berger and Thompson, 1978; Clark and Squire, 1998; Moyer et al, 1990; Solomon et al, 1986; Weiss et al, 1996), the present data are similar to some studies demonstrating normal hippocampal-mediated neuropsychological function in PTSD (Neylan et al, 2004; Pederson et al, 2004; Vasterling et al, 1998)
revious studies that evaluated glucocorticoid sensitivity in PTSD investigated changes in peripheral measures such as salivary cortisol (Goenjian et al, 1996), lymphocytes (Yehuda et al, 1995), mononuclear leukocytes (Yehuda et al, 2004a), plasma immune measures (Rohleder et al, 2004) and dermal vasoconstriction response in the forearm (Coupland et al, 2003)
two tests assay different aspects of hippocampal function. The Wechsler memory scale evaluates declarative memory, while trace eyeblink conditioning evaluates associative learning.
Walter L, Franklin A, Witting A, Wade C, Xie Y, Kunos G, Mackie K, Stella N. 2003. “Nonpsychotropic cannabinoid receptors regulate microglial cell migration” Journal of Neuroscience 23(4): 1398-1405
http://www.ncbi.nlm.nih.gov/pubmed/12598628
pathological overstimulation of neurons by glutamate plus carbachol dramatically increases the production of the endocannabinoid 2-arachidonylglycerol (2-AG) but only slightly increases the production of anandamide
2-AG triggers microglial cell migration. This effect of 2-AG occurs through CB2 and abnormal-cannabidiol-sensitive receptors, with subsequent activation of the extracellular signal-regulated kinase 1/2 signal transduction pathway. It is important to note that cannabinol and cannabidiol, two nonpsychotropic ingredients present in the marijuana plant, prevent the 2-AG-induced cell migration by antagonizing the CB2 and abnormal-cannabidiol-sensitive receptors, respectively
Walter L, Stella N. 2004. “Cannabinoids and neuroinflammation” Br J Pharmacol 141(5): 775-85
http://www.ncbi.nlm.nih.gov/pmc/articles/pmid/14757702/
This review discusses the anti-inflammatory properties of cannabinoid compounds at molecular, cellular and whole animal levels
Ware M, Tongtong W, Shapiro S, Collet J-P. 2015. “Cannabis for the management of pain: assessment of safety study (COMPASS)” The Journal of Pain 16(12): 1233-1242
http://www.jpain.org/article/S1526-5900(15)00837-8/pdf
Two hundred and fifteen individuals with chronic pain were recruited to the cannabis group (141 current users and 58 ex-users) and 216 controls (chronic pain but no current cannabis use) from 7 clinics across Canada. The median daily cannabis dose was 2.5 g/d. There was no difference in risk of serious adverse events (adjusted incidence rate ratio = 1.08, 95% confidence interval = .57–2.04) between groups. Medical cannabis users were at increased risk of non-serious adverse events (adjusted incidence rate ratio = 1.73, 95% confidence interval = 1.41–2.13); most were mild to moderate. There were no differences in secondary safety assessments. Quality-controlled herbal cannabis, when used by patients with experience of cannabis use as part of a monitored treatment program over 1 year, appears to have a reasonable safety profile
The primary outcome consisted of serious adverse events and non-serious adverse events. Secondary safety outcomes included pulmonary and neurocognitive function and standard hematology, biochemistry, renal, liver, and endocrine function
Patients in the cannabis group were younger, with a larger percentage of male, disabled, and tobacco or alcohol users compared with the control group. Socioeconomic status did not differ between the groups. The average pain intensity score at baseline was significantly higher in the cannabis group than in the control group
Sixty-seven patients receiving study cannabis and 34 control patients discontinued the study before the full year of follow-up
Twenty-eight (13%) patients in the cannabis group reported at least 1 SAE, compared with 42 (19%) in the control group. The risk of having at least 1 SAE was not significantly different between the 2 groups (unadjusted OR = .64, 95% CI = .38–1.04) The total number of SAEs was similar in the cannabis and control groups (40 and 56, respectively). The incident rates of SAEs were 22.6 and 27.5 events per 100 person-years of follow-up in the cannabis and control groups, respectively (unadjusted IRR = .82, 95% CI = .46–1.46).
Two participants in the control group died over the course of the trial, 1 by suicide and the other died in the operating room after emergency treatment for abdominal pain; there were no deaths in the cannabis group
Most patients in the cannabis group (190 of 215; 88.4%) and the control group (184 of 216; 85.2%) experienced at least 1 non-serious AE, with a median of 3 events per participant (range = 0–16; interquartile range = 2–5) among cannabis users and a median of 2 events per participant (range = 0–14, interquartile range = 1–4) among controls. The risk of having at least 1 AE did not differ significantly between cannabis users and controls (unadjusted OR = 1.32, 95% CI = .75–2.32) A total of 818 non-serious AEs were reported in the cannabis group, resulting in an incidence rate of 4.61 events/person-year. This rate was significantly higher than in the control group in which there were 581 nonserious AEs and an incidence rate of 2.85 events/ person-year (unadjusted IRR = 1.64, 95% CI = 1.35–1.99)
After adjusting for age, gender, education, alcohol history, disability status, concurrent average pain intensity, quality of life, and clinic sites, no difference in neurocognitive function after 1 year was found between cannabis users and controls
Quality of life: Analysis of the change in the PCS indicated greater improvement of physical function in cannabis users than in controls (2.36 point greater improvement at 6 months, 95% CI = .84–3.88; and 1.62 points at 1 year, 95% CI = .10–3.14)
The sensory component of pain was reduced over 1 year in cannabis users compared with controls (Supplementary Tables 19 and 20). The total symptom distress score of the Edmonton Symptom Assessment Scale was also improved in cannabis users over 1 year (Supplementary Tables 21 and 22). The total mood disturbance scale of the Profile of Mood States showed significant improvement for cannabis users compared with controls, with improvements found in the tension-anxiety, depressiondejection, anger-hostility, and fatigue-inertia subscales
We found 78 respiratory events in the cannabis group and 56 in the control group, and most were considered mild or moderate. No increase in risk of serious respiratory AEs associated with medical cannabis use was detected (1 SAE in the cannabis group, and 7 in the control group)
the rate of non-serious AEs among ‘‘current cannabis users’’ was lower than that among ‘‘ex-cannabis users’’ or ‘‘naive users.’
Given the difference in follow-up (9 visits after baseline in the cannabis group vs 7 in the control group), patients in the cannabis group may have reported AEs otherwise neglected by controls. The effect of this limitation is likely to lead to more exaggerated estimates of AEs among medical cannabis users than among the controls
confounding by indication due to selective prescribing… herbal cannabis was authorized for refractory patients who had more pain and disability than controls
Webb J, Schroeder M, Chee C, Dial D, Hana R, Jefee H, Mays J, Molitor P. 2013. “Left-Handedness among a community sample of psychiatric outpatients suffering from mood and psychotic disorders” Sage Open 3 (4)
http://sgo.sagepub.com/content/3/4/2158244013503166
left-handedness was 11% for mood disorders, which is similar to the rate in the general population. It was 40% in those with psychotic disorders
a study showing that those suffering from schizophrenia who were left-handed were less likely to have a family history of psychosis (Cannon et al., 1995)
Weber K, Rockstroh B, Borgelt J, Awiszus B, Popov T, Hoffmann K, Schonauer K, Watzl H, Propster K. 2008. “Stress load during childhood affects psychopathology in psychiatric patients” BMC Psychiatry 8:63
http://www.ncbi.nlm.nih.gov/pubmed/18651952/
High stress load in childhood and before puberty, but not in adulthood, was related to negative affect in all participants. In patients, high stress load was related to depressive and posttraumatic symptoms, severity of disorder, and the diagnoses of MDD and PD.
Results support the hypothesis of stress-sensitive periods during development, which may interact with genetic and other vulnerability factors in their influence on the progress of psychiatric disorders. A ‘dose’ effect of stress load on the severity of psychopathology is not restricted to the relationship between traumata and PTSD.
Webster P. 2001. “Marijuana and Music: a speculative exploration” Journal of Cannabis Therapeutics 1(2): 93-105
http://www.tandfonline.com/doi/abs/10.1300/J175v01n02_05?journalCode=wzcn20
Weickert C, Fung S, Catts V, Schofield P, Allen K, Moore L, Newell K, Pellen D, Huang X-F, Catts S, Weickert T. 2013. “Molecular evidence of N-methyl-D-aspartate receptor hypofunction in schizophrenia” Mol Psychiatry 18(11): 1185-1192
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3807670/
Modest differences in concentration of NMDAr subunits, misconstrued in the discussion to suggest a much stronger effect than is consistent with the data presented.
Weiland B, Thayer R, Depue B, Sabbineni A, Bryan A, Hutchison K. 2015. “Daily marijuana use is not associated with brain morphometric results in adolescents or adults” Neurobiology of Disease 35(4): 1505-1512
http://www.jneurosci.org/content/35/4/1505.short
Groups were matched on a critical confounding variable, alcohol use, to a far greater degree than in previously published studies. We acquired high-resolution MRI scans, and investigated group differences in gray matter using voxel-based morphometry, surface-based morphometry, and shape analysis in structures suggested to be associated with marijuana use, as follows: the nucleus accumbens, amygdala, hippocampus, and cerebellum. No statistically significant differences were found between daily users and nonusers on volume or shape in the regions of interest. Effect sizes suggest that the failure to find differences was not due to a lack of statistical power, but rather was due to the lack of even a modest effect. In sum, the results indicate that, when carefully controlling for alcohol use, gender, age, and other variables, there is no association between marijuana use and standard volumetric or shape measurements of subcortical structures.
Weinstock M. 2008. “The long-term behavioural consequences of prenatal stress” Neurosci Biobehav Rev 32(6):1073-86
http://www.ncbi.nlm.nih.gov/pubmed/18423592
Maternal distress during pregnancy increases plasma levels of cortisol and corticotrophin releasing hormone in the mother and foetus. These may contribute to insulin resistance and behaviour disorders in their offspring that include attention and learning deficits, generalized anxiety and depression … associated with structural changes in the hippocampus, frontal cortex, amygdala and nucleus accumbens
Whitaker R. 2004. “The case against antipsychotic drugs: a 50-year record of doing more harm than good” Med Hypotheses 62(1): 5-13
http://www.ncbi.nlm.nih.gov/pubmed/14728997
Although the standard of care in developed countries is to maintain schizophrenia patients on neuroleptics, this practice is not supported by the 50-year research record for the drugs
two principles: (a). no immediate neuroleptisation of first-episode patients; (b). every patient stabilized on neuroleptics should be given an opportunity to gradually withdraw from them. This model would dramatically increase recovery rates and decrease the percentage of patients who become chronically ill.
White H, Hansell S. 1998. “Acute and long-term effects of drug use on aggression from adolescence into adulthood” Journal of Drug Issues 28(4): 837-858
http://psycnet.apa.org/psycinfo/1999-00432-002
Heavier marijuana use predicted decreased aggression in later adolescence and adulthood
White H, Bechtold J, Loeber R, Pardini D. 2015. “Divergent marijuana trajectories among men: socioeconomic, relationship, and life satisfaction outcomes in the mid-30s” Drug and Alcohol Dependence 156: 62-69
http://www.drugandalcoholdependence.com/article/S0376-8716(15)01625-7/abstract
Although there were initially group differences across all domains, once pre-existing confounds and co-occurring other substance use were included in the model, groups only differed in terms of partner and friend marijuana use
not at a heightened risk for maladjustment in adulthood except for lower SES among Black men
Whitfield-Gabrieli S, Thermenos H, Milanovic S, Tsuang M, Faraone S, McCarley R, Shenton M, Green A, Nieto-Castanon A, LaViolette P, Wojcik J, Gabrieli J, Seidman L. 2009. “Hyperactivity and hyperconnectivity of the default mode network in schizophrenia and in first-degree relatives of persons with schizophrenia” Proc Natl Acad Sci USA 106(4): 1279-1284
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2633557/
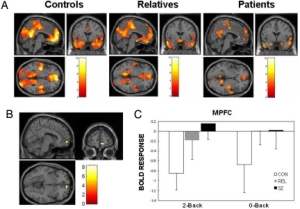
controls exhibited task-related suppression of activation in the default network, including medial prefrontal cortex (MPFC) and posterior cingulate cortex/precuneus. Patients and relatives exhibited significantly reduced task-related suppression in MPFC, and these reductions remained after controlling for performance. Increased task-related MPFC suppression correlated with better WM performance in patients and relatives and with less psychopathology in all 3 groups. For WM task performance, patients and relatives had greater activation in right dorsolateral prefrontal cortex (DLPFC) than controls. During rest and task, patients and relatives exhibited abnormally high functional connectivity within the default network. The magnitudes of default network connectivity during rest and task correlated with psychopathology in the patients. Further, during both rest and task, patients exhibited reduced anticorrelations between MPFC and DLPFC, a region that was hyperactivated by patients and relatives during WM performance. Among patients, the magnitude of MPFC task suppression negatively correlated with default connectivity, suggesting an association between the hyperactivation and hyperconnectivity in schizophrenia. Hyperactivation (reduced task-related suppression) of default regions and hyperconnectivity of the default network may contribute to disturbances of thought in schizophrenia and risk for the illness
10-fold increase in the incidence of the disease in first-degree relatives and a 40–65% concordance rate in identical twins
activation in the default network is associated with spontaneous, internally generated, task-independent mentation, which is greatest during rest
similar pattern in patients and relatives, who were all unmedicated and had never been psychotic, suggests that the findings in patients were not secondary to disease history, which typically includes medication treatment and many psychosocial consequences of schizophrenia, such as social isolation
whereas patients and controls differed by multiple measures in PCC, relatives were similar to controls in PCC by these same measures. This suggests that MPFC dysfunction is related to risk for the disease, whereas PCC dysfunction is related either to greater risk for the disease or to the expression of the illness
hyperactivation and hyperconnectivity of the default network in schizophrenia may constantly divide or misdirect attentional resources when patients attempt to perceive, think about, or act on the external world
among healthy people, lesser suppression of the default network is associated with increased mind-wandering
Constant overengagement of the default network could lead to an exaggerated focus on one’s own thoughts and feelings as well as an ambiguous integration between one’s own thoughts and feelings with events in the environment. Thus, neutral events would seem to be imbued with exaggerated self-relevance, and the boundary between the internal world of reflection and feeling and the external world of perception and action would be weakened
Wigman J, Nierop M, Vollebergh W, Lieb R, Beesdo-Baum, Wittchen H-R, van Os J. 2012. “Evidence that psychotic symptoms are prevalent in disorders of anxiety and depression, impacting on illness onset, risk and severity — implications for diagnosis and ultra-high risk research” Schizophr Bull 38(2): 247-257
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3283146/
Depressive and/or anxiety disorders, with no previous history of psychotic disorder, were examined for the presence of psychotic symptoms in a representative community sample of adolescents and young adults (Early Developmental Stages of Psychopathology study; n = 3021)
Around 27% of those with disorders of anxiety and depression displayed one or more psychotic symptoms, vs 14% in those without these disorders (OR 2.23, 95% CI 1.89–2.66, P < .001). Presence as compared with nonpresence of psychotic symptomatology was associated with younger age (P < .0001), male sex (P< .0058), and poorer illness course (P < .0002). In addition, there was greater persistence of schizotypal (P< .0001) and negative symptoms (P < .0170), more observable illness behavior (P < .0001), greater likelihood of service use (P < .0069), as well as more evidence of familial liability for mental illness (P < .0100), exposure to trauma (P < .0150), recent and more distant life events (P < .0006–.0244), cannabis use (P < .0009), and any drug use (P < .0008)
Copresence of psychotic symptomatology in disorders of anxiety and depression is common and a functionally and etiologically highly relevant feature, reinforcing the view that psychopathology is represented by a network or overlapping and reciprocally impacting dimensional liabilities
Affective dysregulation and reality distortion are correlated but separable dimensions of psychopathology.1The association is present over the continuum of subclinical and clinical expression of psychopathology, although stronger in the clinical range.2 Symptoms and syndromes of depression and anxiety3,4 are present in the majority of patients with schizophrenia, and these affective symptoms may distinguish distinct subgroups within clinical samples of individuals with psychotic illness
genetic studies have suggested familial links between affective and psychotic disorders18,19 as well as between schizophrenia and a range of other mental disorders in the nonpsychotic spectrum,20suggesting shared liabilities
subclinical psychotic experiences predict not only later onset of psychotic disorders23 but also later affective disorders24, even when the psychotic experiences are not considered clinically relevant.25
. Classificatory principles for mental disorders have been dominated by the nomothetic approach in which criteria relating to symptoms and complaints in patients are assumed to be indicators of an underlying latent diagnostic construct. This approach, however, may not be in agreement with evidence that symptoms in practice form part of overlapping and reciprocally impacting dimensional liabilities that give rise to highly patient-specific admixtures and trajectories.29
psychopathology may be considered as a network of symptom dimensions that reciprocally impact each other over time and are linked as part of a homeostatic mechanism, time, and/or share liability
some of the assessments of the exposure variables may have been biased because individuals who are more severely ill may report greater degree of exposure to, eg, childhood trauma or life events
Wilkinson S, Radhakrishnan R, D’Souza D. 2014. “Impact of cannabis use on the development of psychotic disorders” Curr Addict Rep 1(2): 115-128
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4352721/
Exposure to cannabis in adolescence is associated with a risk for later psychotic disorder in adulthood; this association is consistent, temporally related, shows a dose-response, and is biologically plausible. However, cannabis is neither necessary nor sufficient to cause a persistent psychotic disorder. More likely it is a component cause that interacts with other factors to result in psychosis. The link between cannabis and psychosis is moderated by age at onset of cannabis use, childhood abuse and genetic vulnerability
THC produces its psychoactive effects via actions at CB1, where it functions as a partial agonist with modest affinity (Ki = 35-80 nmol) and low intrinsic activity (17). CBD, a major constituent of cannabis that does not produce euphoria, may have anxiolytic and antipsychotic effects in both preclinical and humans studies (reviewed in (18)). The CBD content of cannabis varies and lower levels of CBD in cannabis have been associated with higher rates of psychosis (19–23). For example a variant of South African cannabis that is nearly devoid of CBD is associated with higher rates of psychosis (21, 23, 24). Of note, CBD has been shown to inhibit the psychotomimetic effects of THC (25, 26). Last, it warrants mention that a number of synthetic cannabinoids that are full CB1 agonists with generally higher affinity for CB1 are currently being used by a substantial number of individuals (27)
. In the 19th century, Moreau (de Tours) characterized transient hallucinations, paranoia, dissociative symptoms, thought disorganization and impairments in attention and memory reminiscent of psychotic symptoms seen in schizophrenia in the context of acute cannabis intoxication (reviewed in (28)). These phenomena have also been documented in numerous case-reports (reviewed in (28)) and estimated to occur in about 20%-50% of individuals who use cannabis (29, 30)
Consistent with the acute psychotogenic effects of cannabis, similar psychotic symptoms have been reported with the use of medicinal cannabinoids such as dronabinol, nabilone, and levonantradol (reviewed in (28)(31)). More recently, there is increasing recognition of psychosis related to the recreational use of newer synthetic cannabinoids (32) which are sold as Spice or K2 and which are more potent CB1 agonists than THC(27).
Cannabis, THC and synthetic cannabinoids have been shown to produce a full range of positive symptoms (such as suspiciousness, paranoid and grandiose delusions, conceptual disorganization, fragmented thinking, and perceptual alterations), negative symptoms (such as blunted affect, emotional withdrawal, psychomotor retardation, lack of spontaneity and reduced rapport), and cognitive impairments (such as deficits in verbal learning, short-term memory, working memory, executive function, abstract ability, decision making, attention, and time perception abnormalities) in healthy volunteers that bear resemblance to the symptoms of schizophrenia (25, 33–36). Further, THC exacerbates psychotic symptoms in patients with chronic schizophrenia despite being on stable doses of antipsychotics (37)
THC reduces amplitude of the novelty P300a and target P300b, measures of the automatic orientation of attention (P300a) and context updating (P300b) in healthy participants (38, 39) in a dose-dependent manner (40) without affecting processing speed
Cannabis-induced acute persistent psychosis has been documented in multiple case-series (23, 70–77) characterized by hallucinations, paranoia, delusions, depersonalization, emotional lability, amnesia, confusion and disorientation, which followed the ingestion of large doses of cannabis. These psychotic episodes tend to resolve relatively faster than schizophrenic psychotic episodes, and do not usually recur without re-use of cannabis (23, 74, 77–84) (86). (reviewed in (85)).
It should be noted that most studies have focused on positive symptom outcomes – there is a dearth of studies examining negative symptoms and or cognitive deficits.
the onset of schizophrenia is difficult to establish. The emergence of positive symptoms may be the final step in the evolution of schizophrenia. Negative and cognitive symptoms are more difficult to recognize than positive psychotic symptoms, and often precede positive symptoms. This makes pinpointing the onset of illness and therefore establishing the temporal relationship between cannabis exposure and schizophrenia challenging.
the cannabis-psychosis relationship fulfills many but not all of the traditional criteria for causality
Windle M, Wiesner M. 2004. “Trajectories of marijuana use from adolescence to young adulthood: predictors and outcomes” Dev Psychopathol 16(4): 1007-27
http://www.ncbi.nlm.nih.gov/pubmed/15704825/
five trajectory groups were identified: Abstainers, Experimental Users, Decreasers, Increasers, and High Chronics
High Chronic group, relative to the other trajectory groups, had higher levels of delinquency, lower academic performance, more drug using friends, and more stressful life events
Adolescent trajectory group membership was significantly associated in young adulthood with cannabis and alcohol disorders but not with major depressive disorders or anxiety disorders
van Winkel R, GROUP Investigators. 2011. “Family-based analysis of genetic variation underlying psychosis-inducing effects of cannabis sibling analysis and proband follow-up” Arch Gen Psychiatry 68(2): 148-157
http://archpsyc.jamanetwork.com/article.aspx?articleid=211074
observation of an AKT1 × cannabis interaction. This interaction impacted the short-term psychotomimetic effects of cannabis use in an at-risk population and, in addition, also influenced long-term developmental effects on psychotic disorder. AKT1 is a serine/threonine kinase that is activated by phosphatidylinositol-3-kinase (PI3K).38 One of the essential functions of AKT is the phosphorylation of glycogen synthase kinase (GSK-3) at Ser21 in GSK-3α and Ser9 in GSK-3β, causing its inactivation.39 AKT and GSK-3 have emerged as the focal point for many signal-transduction pathways, regulating multiple cellular processes including transcription, apoptosis, endoplasmic reticulum stress response, cell proliferation, and cell survival.38 Importantly, cannabinoids are able to activate the AKT1/PI3K pathway by acting on CB1 and CB2 receptors in vitro.40 Moreover, immediate administration of THC in mice activates AKT1 in vivo (through AKT1 phosphorylation) in several brain areas, including the striatum, independent of dopaminergic D1 and D2 receptor blockade.41
Decreased AKT1 levels have been observed in lymphoblasts and the postmortem prefrontal cortex of patients with schizophrenia,42,43 and several studies have shown evidence for genetic association with schizophrenia,43– 47 although not all studies were able to confirm this.48,49
dopamine D2 receptors may signal through an AKT1/GSK-3 signaling pathway via β-arrestin 2, and multiple lines of evidence support the involvement of the β-arrestin-2/AKT/GSK-3 pathway in the regulation of dopamine-associated behaviors and the response to antipsychotic treatment.52 If psychotomimetic effects of THC are indeed modulated by the AKT1 /GSK-3 signaling cascade, this could potentially explain why dopamine D2 receptor blockade is ineffective in reducing psychotomimetic effects of THC in healthy individuals53
the hypothesized cannabinoid-regulated AKT1/GSK modulation would occur downstream of the dopamine D2 receptor
in contrast to previous studies, found no evidence that SNPs in COMT interact with cannabis use to influence positive schizotypy in unaffected siblings
NRG1-induced AKT1 phosphorylation was significantly diminished in COMT Val carriers in both normal subjects and in patients with schizophrenia in this cell model, as was NRG1-induced translocation of AKT1 to the plasma membrane.56 These findings suggest that the previously reported COMT × cannabis findings and the present finding of AKT1 × cannabis interaction may represent different genetic signals pointing to the same, or related, underlying molecular mechanism
Witkin J, Tzavara E, Nomikos G. 2005. “A role for cannabinoid CB1 receptors in mood and anxiety disorders” Behavioural Pharmacology 16:315-331
http://www.ncbi.nlm.nih.gov/pubmed/16148437
fuzzy picture, anxiolytic and anxiogenic. Reviews ECS alterations in various disorders, gushes about the possible uses of Rimonabant and FAAH inhibitors
offer the possibility of pharmacotherapy of anxiety disorders through enhanced, ‘use-dependent’ stimulation of CB1 receptors, avoiding the overt and indiscriminate effects, neuroadaptive processes that result from use of an in direct agonist
suggest a role of compounds that enhance endocannabinoid neurotransmission in counteracting impulsivity associated with diverse neuropsychiatric ailments
Jin et al. (2004) have shown a CB1 receptor-independent but TRPV1 receptor-dependent neuro-genesis-promoting effect of rimonabant in KO mice
antagonist induced improvement in retention is accompanied by deficits in reversal learning, evidenced as increased perseverations in the Morris water maze (Varvel and Lichtman, 2002)
Improved long-term memory, but cognitive inflexibility, due possibly to proactive interference, was also demonstrated in chick-adees after CB1 receptor antagonist administration (Shiflett et al. , 2004)
Wittchen H-U, Fröhlich C, Behrendt S, Günther A, Rhm J, Zimmerman P, Lieb R, Perkonigg A. 2007. “Cannabis use and cannabis use disorders and their relationship to mental disorders: A 10-year prospective-longitudinal community study in adolescents.” Drug and Alcohol Dependence 88(1):S60-S70
http://www.sciencedirect.com/science/article/pii/S0376871606004844
A range of psychopathological conditions, including depressive, bipolar and less consistently anxiety disorders as well as the degree of their comorbidity are significantly associated with incident CU and progression to CUD, even when controlling for externalising disorders. A better understanding of this complex interplay may result in better aetiological models and intervention strategies.
Wolf S, Tauber S, Ullrich O. 2008. “CNS immune surveillance and neuroinflammation: endocannabinoids keep control” Curr Pharm Des 14(23): 2266-78
http://www.ncbi.nlm.nih.gov/pubmed/18781977
The immune system is allowed to keep the CNS under surveillance, but in a strictly controlled, limited and well-regulated manner. The first line of defense lies outside the brain parenchyma to spare neuronal tissue from the detrimental effects of an inflammatory immune response. As a second line of defense neuroinflammation is unavoidable when pathogens infiltrate the brain or the CNS-immune-homeostasis fails. Inflammation in the CNS is often accompanied by divers brain pathologies
the endocannabinoid system participates crucially in the immune control and protection of the CNS
Wolff M, Leander J. 2003. “SR141716A, a cannabinoid CB1 receptor antagonist, improves memory in a delayed radial maze task” Eur J Pharmacol 477(3):213-7
http://www.ncbi.nlm.nih.gov/pubmed/14522359
Although SR141716A had no effect on the number of errors committed after a short interval, SR141716A significantly reduced the number of errors that occurred after 7 h
Woodberry K, Giuliano A, Seidman L. 2008. “Premorbid IQ in Schizophrenia: A Meta-Analytic Review” American Journal of Psychiatry 165(5): 579-587
http://psychiatryonline.org/doi/abs/10.1176/appi.ajp.2008.07081242?url_ver=Z39.88-2003
Years before the onset of psychotic symptoms, individuals with schizophrenia, as a group, demonstrate mean IQ scores approximately one-half of a standard deviation below that of healthy comparison subjects
Woon F. Sood S. Hedges D. 2010. “Hippocampal volume deficits associated with exposure to psychological trauma and posttraumatic stress disorder in adults: a meta-analysis” Prog Neuropsychopharmacol Biol Psychiatry 34(7): 1181-8
http://www.ncbi.nlm.nih.gov/pubmed/20600466
We identified 39 hippocampal volumetric studies in adults with PTSD compared to control groups consisting of either trauma-exposed controls without PTSD or trauma-unexposed controls, or both
Hippocampal volumes were smaller in the PTSD group and trauma-exposed group without PTSD compared to the trauma-unexposed group. Further, the right hippocampus was smaller in the PTSD group compared to the trauma-exposed group without PTSD. Additionally, the right hippocampus was larger than the left in the PTSD and trauma-unexposed groups but not in the trauma-exposed group without PTSD. Hippocampal volume reduction is associated with trauma exposure independent of PTSD diagnosis, albeit additional hippocampal reduction was found in PTSD compared to the trauma-exposed group without PTSD.
Wright R, Conrad C. 2008. “Enriched environment prevents chronic stress-induced spatial learning and memory deficits” Behav Brain Res 187(1): 41-47
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2629380/
Chronic stress impairs spatial memory and alters hippocampal structure, which are changed in the opposite direction following enriched environment (EE)
EE in adulthood prevented spatial learning and memory impairment in chronically stressed rats, indicating that the context of stress exposure impacts susceptibility to chronic stress-induced cognitive deficits
retraction of the apical dendrites in the CA3 region of the hippocampus following prolonged restraint [9, 17, 33] or chronic social stress [18]. Chronic stress can also modify hippocampal dendritic spine number and shape [11, 19, 30]. Prolonged stressful periods can result in cell death [31]. Collectively there is clear evidence that chronic stress can significantly alter hippocampal structure
Chronic stress was perceived as stressful because STR-STD showed reduced weight gain, and reduced body weight in the STR-EE group began before the onset of restraint
EE can increase hippocampal neurogenesis [6], dendritic spines [4] dendritic arborization [12], BDNF [26], and glucocorticoid receptors [20] … EE may also prevent chronic stress-induced spatial learning and memory impairments through alterations of the HPA axis
Wu N, Schairer L, Dellor E, Grella C. 2010. “Childhood trauma and health oucomes in adults with comorbid substance abuse and mental health disorders” Addict Behav 35(1): 68-71
http://www.ncbi.nlm.nih.gov/pubmed/19775820/
Adults with SUDs/MHPs (N=402) were recruited from residential treatment programs and interviewed at treatment admission. Exposures to 9 types of adverse childhood experiences were summed and categorized into 6 ordinal levels of exposure… Most of the sample reported exposure to CTEs, with higher exposure rates among the study sample compared with the primary health care sample. Greater exposure to CTEs significantly increased the odds of several adverse adult outcomes, including PTSD, alcohol dependence, injection drug use, tobacco use, sex work, medical problems, and poor quality of life. Study findings support the importance of early prevention and intervention and provision of trauma treatment for individuals with SUDs/MHPs
Wunderink L, Nieboer R, Wiersma D, Sytema S, Nienhuis F. 2013. “Recovery in remitted first-episode psychosis at 7 years of follow-up of an early dose reduction/discontinuation or maintenance treatment strategy” JAMA Psychiatry 70(9): 913-920
http://archpsyc.jamanetwork.com/article.aspx?articleid=1707650
The drug reduction patients experienced twice the recovery rate of the maintenance treatment (40.4% vs 17.6%). Logistic regression showed an odds ratio of 3.49 (P = .01). Better DR recovery rates were related to higher functional remission rates in the DR group but were not related to symptomatic remission rates
those who stayed on antipsychotics had worse outcomes
X M, Haley A. 1965. The Autobiography of Malcolm X Grove Press
http://al-rasid.com/shared_uploads/The.Autobiography.of.MalcolmX.pdf
“I have no mercy or compassion in me for a society that will crush people, and then penalise them for not being able to stand up under the weight”
his mother became schizophrenic after years of trauma and racist abuse. He was also beaten as a child and suffered many traumas, used drugs extremely heavily (including marijuana “almost by the ounce”), sentenced to 10 years in prison for armed robbery, provides ample harsh criticism of the social injustices which made criminality inevitable for him and his peers. His brother Reginald also became schizophrenic after an unexpected social exclusion from his family and religion.
Xiong W, Cheng K, Cui T, Godlewski G, Rice K, Xu Y, Zhang L. 2011. “Cannabinoid potentiation of glycine receptors contributes to cannabis-induced analgesia” Nat Chem Biol 7(5): 296-303
http://www.ncbi.nlm.nih.gov/pmc/articles/pmid/21460829/
Yang C, Huang C, Hsu K. 2005. “Behavioral stress enhances hippocampal CA1 long-term depression through blockade of the glutamate uptake” J Neurosci 25(17): 4288-93
http://www.ncbi.nlm.nih.gov/pubmed/15858055
We also found that stress induces a profound decrease in the glutamate uptake in the synaptosomal fraction of the hippocampal CA1 region
Yehuda R. 2002. “Post-Traumatic Stress Disorder” N Engl J Med 346: 108-114
http://www.nejm.org/doi/full/10.1056/NEJMra012941
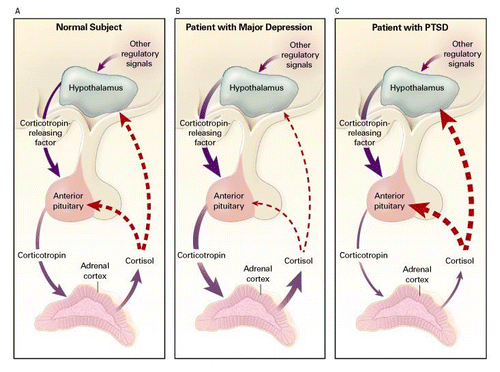
Yehuda R, Golier J, Kaufman S. 2005. “Circadian rhythm of salivary cortisol in Holocaust survivors with and without PTSD” Am J Psychiatry 162(5): 998-1000
http://www.ncbi.nlm.nih.gov/pubmed/15863805/
Cortisol levels were significantly lower at awakening, at 8:00 a.m., and at 8:00 p.m. in Holocaust survivors with PTSD than in nonexposed subjects, resulting in a flatter circadian rhythm, similar to what has been observed in aging but different from what has been reported in younger subjects with PTSD.
Young M, Read J, Barker-Collo S, Harrison R. 2001. “Evaluating and overcoming barriers to taking abuse histories” Professional Psychology: Research and Practice 32(4): 407-414
http://psycnet.apa.org/index.cfm?fa=buy.optionToBuy&id=2001-07733-012
Psychologists and psychiatrists often resistant to taking abuse histories. Reasons include more pressing issues, fear of disturbing clients, a diagnosis of schizophrenia, biological etiology beliefs, and fear of inducing “false memories.”
Yuan M, Kiertscher S, Cheng Q, Zoumalan R, Tashkin D, Roth M. 2002. “Delta 9-Tetrahydrocannabinol regulates Th1/Th2 cytokine balance in activated human T cells” J Neuroimmunol 133(1-2): 124-31
http://www.ncbi.nlm.nih.gov/pubmed/12446015
THC suppressed T cell proliferation, inhibited the production of interferon-gamma and shifted the balance of T helper 1 (Th1)/T helper 2 (Th2) cytokines. Intracellular cytokine staining demonstrated that THC reduced both the percentage and mean fluorescence intensity of activated T cells capable of producing interferon-gamma, with variable effects on the number of T cells capable of producing interleukin-4. Exposure to THC also decreased steady-state levels of mRNA encoding for Th1 cytokines, while increasing mRNA levels for Th2 cytokines. The CB2 receptor antagonist, SR144528, abrogated the majority of these effects. We conclude that cannabinoids have the potential to regulate the activation and balance of human Th1/Th2 cells by a CB2 receptor-dependent pathway
Yücel M, Solowij N, Respondek C, Whittle S, Fornito A, Pantelis C, Lubman DI. 2008. “Regional brain abnormalities associated with long-term heavy cannabis use.” Arch Gen Psychiatry 65(6): 694-701
http://www.ncbi.nlm.nih.gov/pubmed/18519827/
Fifteen carefully selected long-term (>10 years) and heavy (>5 joints daily) cannabis-using men
Cannabis users had bilaterally reduced hippocampal and amygdala volumes (P = .001), with a relatively (and significantly [P = .02]) greater magnitude of reduction in the former (12.0% vs 7.1%). Left hemisphere hippocampal volume was inversely associated with cumulative exposure to cannabis during the previous 10 years (P = .01) and subthreshold positive psychotic symptoms
Yung A, Yuen H, Berger G, Francey S, Hung T-C, Nelson B, Phillips L, McGorry P. 2007. “Declining transition rate in ultra high risk (prodromal) services: dilution or reduction of risk?” Schizophr Bull 33(3): 673-681
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2526154/
There is growing interest in the prodromal stage of psychotic disorders, with many services now providing care for these “ultra high risk” (UHR) individuals. However, a reduction in the rate of transition to psychosis has been suspected over the last few years… each successive year showing a rate 0.80 times that of the preceding year. Functioning and symptom level were not responsible for the change. The decreased transition rate was partly explained by a reduction in the duration of symptoms of patients prior to receiving help. That is, UHR individuals are being detected and provided with care earlier than in the past. Thus, the decline in transition rate may be due to treatment being more effective at this very early stage of illness or it may be due to finding more false positives who were never at risk of psychosis, ie, a “dilution” effect
3 criteria are currently used to identify the UHR group.12,13 These are (1) Attenuated Psychotic Symptoms (APS) Group: have experienced subthreshold, attenuated positive psychotic symptoms during the past year; (2) Brief Limited Intermittent Psychotic Symptoms (BLIPS) Group: have experienced episodes of frank psychotic symptoms that have not lasted longer than a week and have been spontaneously abated; or (3) Trait plus State Risk Factor Group: have a first-degree relative with a psychotic disorder or the identified client has a schizotypal personality disorder and they have experienced a significant decrease in functioning during the previous year. Additionally, a perceived need for psychiatric help is also required as the person needs to have been referred to a specialized psychiatric service for management, the Personal Assessment and Crisis Evaluation (PACE) Clinic
Using these UHR criteria, we found a rate of transition to psychosis of over 34% within 6 months of referral to the PACE service and between 35% and 40% within 12 months.12,13 These rates are several thousand fold over the expected incidence rate for first-episode psychosis in the general population
Recent data from the PACE Clinic found a 6-month transition rate of only 9.2%.2
Psychosocial treatment may be harmful too, if it involves educating individuals about psychosis and informing them that they are at risk, if they are not actually at risk. Treatment may needlessly alarm them, result in possible curtailment of activities, and cause stigma including self-stigmatization (eg, they may decide to cease attending university or work to avoid stress).23–25
we are detecting UHR individuals and providing them with care much earlier than in the past
Yung A, Nelson B, Stanford C, Simmons M, Cosgrave E, Killackey E, Phillips L, Bechdolf A, Bucky J, McGorry P. 2008. “Validation of ‘prodromal’ criteria to detect individuals at ultra high risk of psychosis: 2 year follow up” Schizophr Res 105(1-3): 10-7
http://www.ncbi.nlm.nih.gov/pubmed/18765167
The criteria significantly predicted onset of psychotic disorder within 2 years. The transition rate of 16% was much lower than in initial cohorts (over 40%).
Yung A, Cotter J, Wood S, McGorry P, Thompson A, Nelson B, Lin A. 2015. “Childhood maltreatment and transition to psychotic disorder independently predict long-term functioning in young people at ultra-high risk for psychosis” Psychological Medicine 45(16): 3453-3465
http://journals.cambridge.org/action/displayAbstract?fromPage=online&aid=10017640&fulltextType=RA&fileId=S003329171500135X
Participants were 268 (152 females, 116 males) individuals identified as UHR 2–14 years previously
Baseline negative symptoms, impaired emotional functioning, disorders of thought content, low functioning, past substance use disorder and history of childhood maltreatment predicted poor functioning at follow-up in univariate analyses. Only childhood maltreatment remained significant in the multivariate analysis (p < 0.001)
Childhood maltreatment was a significant predictor of poor functioning in both the transitioned and non-transitioned groups
Zalesky A, Solowij N, Yucel M, Lubman D, Takagi M, Harding I, Lorenzetti V, Wang R, Searle K, Pantelis C, Seal M. 2012. “Effect of long-term cannabis use on axonal fibre connectivity” Brain 2012: 135; 2245-2255
http://www.ncbi.nlm.nih.gov/pubmed/22669080 in dropbox
Axonal connectivity impaired in the right fimbria of the hippocampus (fornix), splenium of the corpus callosum and commissural fibres. Radial and axial diffusivity associated with the age at which regular cannabis use commenced.
In adulthood, density of cannabinoid receptors in these structures diminishes (Romero et al ., 1997; Glass et al ., 1998)
Users had greater trait anxiety (State-Trait Anxiety Inventory) and depressive symptoms (Beck Depression Inventory) and lower Global Assessment of Functioning scores.
No between-group differences were found in the number of streamlines comprising any of the 20 bins, no evidence of any brain-wide shortening of axonal fibres. No between-group difference was found in whole-brain volume
commissural fibre beginning at the splenium of the corpus callosum and extending deep within the right precuneus. … a total of 9 1.7 streamlines in users and 78 15.1 in non-users, an 88% reduction.
fimbria can be seen to extend medially towards the hippocampal commissure, not into left hemisphere. fimbria network comprised a total of 7 1.6 streamlines in users (mean standard error) and 44 6.4 in non-users, an 84% reduction
No associations with duration of regular cannabis use and cumulative life dose were found. No association was found between the duration of regular cannabis use and the age at which regular cannabis use commenced.
Zammit S, Allebeck P, Lewis G. 2002. “Self reported cannabis use as a risk factor for schizophrenia in Swedish conscripts of 1969: historical cohort study” BMJ 325(7374): 1199
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC135490/
psychiatric diagnosis at conscription, IQ score, personality variables concerned with interpersonal relationships, place of upbringing, paternal age, and cigarette smoking are all associated with schizophrenia.10–13,20,21 We included these variables as potential confounders in the regression model. Disturbed behaviour in childhood, history of alcohol misuse, family history of psychiatric illness, financial situation of the family, and father’s occupation were also considered to be potential confounders and included in the analysis
Out of the 50053 subjects, 362 received diagnosis of schizophrenia by 1996
5391 subjects (10.8% of the cohort) had ever used cannabis, and 73 of these (1.4%) developed schizophrenia. In 69 subjects who started using drugs before 1969, 19 (31%; 95% confidence interval 20% to 44%) of those developing schizophrenia had stopped using drugs before conscription, as opposed to 2810 (64%; 62% to 65%) of the 4418 who did not develop schizophrenia (P<0.001)
adjusted odds ratio for subjects with a history of heaviest use of cannabis (>50 occasions) was 3.1
Subjects who had ever used cannabis had an increased risk of developing a psychosis other than schizophrenia (crude odds ratio 1.4, 1.1 to 1.9), but this effect was reduced after adjustment (adjusted odds ratio 1.1, 0.8 to 1.5) A similar pattern was observed for the association with cannabis frequency, with a linear trend odds ratio of 1.1 (1.0 to 1.2, P<0.02) before adjustment and of 1.0 (0.9 to 1.1, P<0.85) after adjustment
For all the analyses, diagnosis on conscription, IQ score, and place of upbringing contributed most to confounding
no association between cannabis and other psychotic illnesses, which implies that cannabis has a rather specific association with an increased risk of schizophrenia
the association between cannabis and schizophrenia persisted even after adjusting for use of alcohol, cigarettes, and other drugs
Zarate C, Singh J, Carlson P, Brutsche N, Ameli R, Luckenbaugh D, Charney D, Manji H. 2006. “A randomized trial of an n-methyl-d-aspartate antagonist in treatment resistant major depression” Arch Gen Psychiatry 63(8): 856-864
http://archpsyc.jamanetwork.com/article.aspx?articleid=668195
Zarate C, Brutsche N, Ibrahim L, Franco-Chaves J, Diazgranados N, Cravchik A, Selter J, Marquardt C, Liberty V, Luckenbaugh D. 2012. “Replication of ketamine’s antidepressant efficacy in bipolar depression: a randomized controlled add-on trial” Biological Psychiatry 71(11): 939-946
http://www.biologicalpsychiatryjournal.com/article/S0006-3223(11)01210-8/abstract
Zavitsanou K, Garrick T, Huang X. 2004. “Selective antagonist [3H]SR141716A binding to cannabinoid CB1 receptors is increased in the anterior cingulate cortex in schizophrenia” Prog Neuropsychopharmacol Biol Psychiatry 28(2):355-60
http://www.ncbi.nlm.nih.gov/pubmed/14751433
A significant 64% increase in [3H]SR141716A specific binding to CB1 receptors was found in the schizophrenia group as compared to the control group (mean+/-S.E.M.: 46.15+/-6.22 versus 28.02+/-4.20 fmol/mg estimated tissue equivalents; p=0.03). The present results support the suggestion that changes in the endogenous cannabinoid system in the ACC may be involved in the pathology of schizophrenia particularly in relation to negative symptoms
Zhou Y, Liang M, Tian L, Wang K, Hao Y, Liu H, Liu Z, Jiang T. 2007. “Functional disintegration in paranoid schizophrenia using resting-state fMRI” Schizophrenia Research 97: 194-205
http://www.ncbi.nlm.nih.gov/pubmed/17628434 in dropbox
Compared to healthy volunteers, patients showed significant differences in connectivity within networks and between networks, most notably in the connectivities associated with the bilateral dorsal medial prefrontal cortex, the lateral parietal region, the inferior temporal gyrus of the “task-negative” network and with the right dorsolateral prefrontal cortex and the right dorsal premotor cortex of the “task-positive ” network… could be the source of abnormalities in the coordination of and competition between information processing activities in the resting brain of paranoid patients
attention deficit and deficits in monitoring one’s own and other people’s thoughts and intentions (mentalizing) are often observed in patients with schizophrenia (Brune, 2005; Malaspina
et al., 2004; Nuechterlein et al., 2004)… may be related to a failure to integrate the activity of local and distributed neural circuits (Andreasen, 1999; Benes, 2000; Bullmore, 1997; Friston, 1998; Lewis and Gonzalez-Burgos, 2000; Selemon and Goldman-Rakic, 1999; Setephan, 2006)… evidence… widely distributed functional disintegration such as fronto-temporal (Lawrie, 2002; Spence, 2000; Stephan, 2006) fronto-parietal (Kim, 2003), dorsolateral prefrontal-hippocampal formation dysconnection (Meyer-Lindenberg, 2005), fuctional disintegration in cortico-cerebellar-thalamo-cortical circuit (Schlosser, 2003)
unlike previous studies on the resting brain function in schizophrenia (Liang, 2006; Liu, 2006), the present study focused on the intrinsic organization of brain function… abnormal correlations/anti-correlations between regions were widely distributed either within or between networks in patients with paranoid schizophrenia… suggested a source for the abnormalities in coordination of and competition between information processing activities in resting brain of paranoid patients… connectivity within networks primarily increased… implying a concomitant increase in synchrony within either network… increased functional connectivities (task-positive) associated with the R.DLPFC, SMA and bilateral OFG may be related to excessive alertness or sensitivity to external environment… within the task-negative network, increased connectivities associated with bilateral dMPFC, ITG, left LP, and PCC/PCu… associated with mentalizing
abnormal anti-correlations associated with bilateral dMPFC, LP and ITG of the “task negative” network and the R.DLPFC, bilateral insula and OFG of “task positive”… compete in the resting brain of healthy subjects but are balanced… increase in anti-correlations in paranoid patients may suggest excessive competition, contributing to over-mentalizing and attention deficit
all decreased anti-correlations were related to the R.dPM.
abnormal increased synchrony may be explained as resulting from cortical hyperexcitability or abnormally increased connectivity (excessive or erroneous synaptic connectivity)
improper functional integration in schizophrenia not only indicates less interaction between neural units, but also implies the possibility of excessive enhanced interaction. Overconnectivity (overintegration) and disconnectivity (disintegration) between and within interconnected systems may result in different profiles of schizophrenic symptoms (Peled, 1999)
Zhu W, Friedman H, Klein T. 1998. “Δ9-Tetrahydrocannabinol Induces Apoptosis in Macrophages and Lymphocytes: Involvement of Bcl-2 and Caspase-1” JPET 286(2): 1103-1109
http://jpet.aspetjournals.org/content/286/2/1103.long
DNA fragmentation and cell membrane integrity (assessed by dye exclusion) showed that fragmentation preceded membrane damage indicating that THC induced apoptosis rather than cell necrosis. In addition, THC treatment of splenocytes resulted in a decrease of Bcl-2 mRNA and protein as measured by Northern and Western blotting, respectively, and the drug induced apoptosis was blocked by the caspase inhibitor, Ac-Tyr-Val-Ala-L-aspartic acid aldehyde. These data suggest that THC treatment of cultured immune cells induces apoptosis through the regulation of Bcl-2 and caspase activity
Zhu P, Lovinger D. 2005. “Retrograde endocannabinoid signaling in a postsynaptic neuron/synaptic bouton preparation from basolateral amygdala” J Neurosci 25(26): 6199-6207
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1352167/
A CB1 agonist decreased sIPSC frequency and amplitude, whereas CB1 antagonists increased these responses
The BLA has a critical role in fear conditioning (Davis et al., 1994) and appears to provide the anatomical substrate for fear memory storage (LaLumiere et al., 2003; Gale et al., 2004). GABAergic control of projection neuron activity is thought to play a crucial role in information processing in the amygdala (Lang and Paré, 1998). CB1 immunoreactivity was detected in a distinct subset of GABA-containing interneurons in the BLA (Katona et al., 2001), the nucleus in which aversive memories might be formed and stored (Medina et al., 2002). Furthermore, endocannabinoid signaling in BLA has been implicated in extinction of aversive memories (Marsicano et al., 2002)
Ziring D, Wei B, Velazquez P, Schrage M, Buckley N, Braun J. 2006. “Formation of B and T cell subsets require the cannabinoid receptor CB2” Immunogenetics 58(9): 714-25
http://www.ncbi.nlm.nih.gov/pubmed/16924491/
mice deficient in CB2, the Gi-coupled peripheral endocannabinoid receptor, have profound deficiencies in splenic marginal zone, peritoneal B1a cells, splenic memory CD4+ T cells, and intestinal natural killer cells and natural killer T cells
endocannabinoid system is required for the formation of T and B cell subsets involved in immune homeostasis
Because levels of endocannabinoids are highly restricted microanatomically, local regulation of their production and receptor expression offers a new principle for regional immune homeostasis and disease susceptibility
Zoppi S, Perez- Nievas B, Madrigal J, Manzanares J, Leza J, Garcia-Bueno B. 2011. “Regulatory role of cannabinoid receptor 1 in stress-induced excitotoxicity and neuroinflammation” Neuropsychopharmacology 36(4): 805-818
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3055736/
Exposure to stress elicits excitoxicity and neuroinflammation in the brain, contributing to cell death and damage in stress-related neurological and neuropsychiatric diseases
Stress exposure increased CB1 mRNA and protein expression in the prefrontal cortex of WT mice in a mechanism related to N-methyl–aspartate glutamate receptor activation. Daily ACEA pretreatment prevented stress-induced: (1) upregulation of CB1 mRNA and protein, (2) decrease in glutamate uptake and glutamate astroglial transporter excitatory amino acid transporter 2 expression, (3) increase in consecutive proinflammatory molecules, such as cytokines (tumor necrosis factor-α and MCP-1), nuclear factor kappa B, and enzymatic sources, such as inducible nitric oxide synthase (NOS-2) and cyclooxygenase-2 (COX-2), (4) increase in lipid peroxidation; although having no effect on plasma corticosterone. Interestingly, a possible related mechanism could be the positive ACEA modulation of the antiinflammatory pathway deoxyprostaglandin/peroxisome proliferator-activated receptor γ (15d-PGJ2/PPARγ)
lack of CB1 produces hypothalamic/pituitary/adrenal (HPA) axis dysregulation and exacerbates stress-induced excitotoxic/neuroinflammatory responses. These multifaceted neuroprotective effects suggest that CB1 activation could be a new therapeutic strategy against neurological/neuropsychiatric pathologies with HPA axis dysregulation and an excitotoxic/neuroinflammatory component in their pathophysiology
stress exposure elicits excitotoxicity due to the massive release of the excitatory amino acid, glutamate, in some brain areas (eg, the prefrontal cortex) (Moghaddam, 1993), inducing the release of proinflammatory cytokines such as tumor necrosis factor-α(TNF-α) or interleukin 1β (Madrigal et al, 2002). Stress also activates the nuclear transcription factor-kappa B (NF-κB) pathway in a TNF-α-dependent mechanism (Bierhaus et al, 2003; Madrigal et al, 2002). NF-κB activation elicits the expression and activity of proinflammatory enzymatic sources, such as inducible nitric oxide (NO) synthase (NOS-2), and cyclooxygenase-2 (COX-2), among others (Madrigal et al, 2001a, 2003). The result of this sequence of events is the accumulation of oxidative and nitrosative mediators, which can attack membrane phospholipids and cause cell damage in a process known as lipid peroxidation (McEwen, 1998; Madrigal et al, 2001b)
currently, the endocannabinoid system (ECS) is considered as an endogenous homeostatic system activated by different immune challenges, restoring brain balance at different levels (Mechoulam and Shohami, 2007) in different experimental settings (Galve-Roperh et al, 2008; Wolf et al, 2008; Bambico et al, 2009; Cabral and Griffin-Thomas, 2009)
the ECS in stress-responsive neural circuits suggests that it may have a critical role in regulating hypothalamic/pituitary/adrenal (HPA) axis activation and the resulting excitotoxic/neuroinflammatory response… pharmacological enhancement of CB1 protects neurons from acute brain injury
Q-PCR studies showed a consistent increase in TNF-α mRNA in WT mouse prefrontal cortex after stress, an effect that is blocked by ACEA (). Conversely, stressed CB1 KO animals presented a reliable increase in the mRNA of the proinflammatory cytokine, suggesting a potentially deleterious chronic inflammatory status in brain. Stress also increased MCP-1 levels in the prefrontal cortex of WT mice and pharmacological stimulation of CB1 decreased them. CB1 KO mice showed no differences from WT in nonstressed animals. However, stress increased MCP-1 levels in brain in SKO group compared with SWT group
CB1 receptor activation with ACEA was able to completely block NOS-2 upregulation produced by stress exposure ()
one candidate of interest is the antiinflammatory pathway L-PGDS/15d-PGJ2/PPARγ, that is activated after inflammatory/immune stimuli (stress included) and whose activation (even by some cannabinoids) has been shown to confer neuroprotection after stress exposure, and in diverse CNS pathologies with an inflammatory component in their pathophysiology
Stress exposure is a major contributing factor to cell death and damage in neurological and neuropsychiatric diseases by three interrelated mechanisms: HPA axis dysregulation, excitotoxicity, and neuroinflammation. However, stress exposure also activates alternative mechanisms aimed at correctly resolving this response. In this way, the CB1 upregulation demonstrated here could be a mechanism related to this protective response of the brain. Similarly, endocannabinoids are released to negatively regulate HPA axis activation (Manzanares et al, 1999; Patel et al, 2004), and brain SEC activation has been considered as a protective mechanism aimed at reducing both neurodegenerative and inflammatory damage after an immune/inflammatory challenge (Centonze et al, 2008)
our data are in agreement with several studies that propose a role for CB1 as a main regulator of glutamate neurotransmission and metabolism in multiple pathological scenarios (Parmentier-Batteur et al, 2002; Marsicano et al, 2003; Veldhuis et al, 2003; Fernández-López et al, 2006; Kim et al, 2006;Gilbert et al, 2007; Loría et al, 2010)
CB1 receptor activation with ACEA enhances the main glutamate uptake mechanism, by restoring protein expression of the glutamate transporter EAAT-2. Astrocyte uptake of glutamate is a high-affinity process regulated by EAAT-2 (Hertzet al, 1983); the fact that stress decreases EAAT-2 expression in the cerebral cortex (García-Bueno et al, 2007), indicates a specific damaging effect at this level, and makes the effect of ACEA more interesting. CB1 effects on EAAT-2 have also been demonstrated in other experimental settings, such as an in vitromodel of multiple sclerosis (MS) (Loría et al, 2010), or after prenatal exposure to WIN 55,212-2 (Castaldoet al, 2007)
Two main NF-κB-dependent enzymatic sources of oxidative/nitrosative mediators after stress in the brain are NOS-2 and COX-2. NOS-2 expression is downregulated by selective CB1 activation in neurons and astrocytes both in in vitro and in vivo inflammatory-related models (Fernández-López et al, 2006; Espositoet al, 2006; Sheng et al, 2005)… However, cannabinoid–cyclooxygenase interactions may be more intricate
Zuardi A, Crippa J, Hallak J, Moreira F, Guimaraes F. 2006. “Cannabidiol, a Cannabis sativa constituent, as an antipsychotic drug” Braz J Med Biol Res 39(4): 421-9
http://www.ncbi.nlm.nih.gov/pubmed/16612464
The results of two studies on healthy volunteers using perception of binocular depth inversion and ketamine-induced psychotic symptoms supported the proposal of the antipsychotic-like properties of CBD. In addition, open case reports of schizophrenic patients treated with CBD and a preliminary report of a controlled clinical trial comparing CBD with an atypical antipsychotic drug have confirmed that this cannabinoid can be a safe and well-tolerated alternative treatment for schizophrenia.